

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Diabetes mellitus is a serious metabolic disease, and the number of people with diabetes is rapidly increasing due to population growth, aging, urbanization, and increasing prevalence of obesity.1,2 Hyperglycaemia-induced oxidative stress, as a result of excessive production of reactive oxygen species (ROS), plays a key role in the pathogenesis of diabetes-related complications.3-5 Theoretically, suppression of oxidative stress using antioxidants should reverse adverse effects induced by hyperglycaemia in diabetic patients. However, large randomized clinical trials have failed to provide convincing evidence for effective antioxidant therapy with classic antioxidants, such as vitamins E and C.6-8 Therefore, development of new therapeutic regimes is important for patients with diabetes.
Oxidative stress results from increased production of ROS or diminished antioxidant capacity in cells or tissues. One of the major sources of ROS in tissues of diabetic patients is nicotinamide adenine dinucleotide phosphate (NADPH) oxidase.9,10 NADPH oxidase contains two membrane-bound subunits gp91phox (Nox2) and p22phox and cytoplasmic subunits such as p47phox, p67phox and a low-molecular-weight G protein (rac 1 and rac 2).11 NADPH oxidase catalyzes the generation of superoxide anion using NADPH as the electron donor in cells.12 Superoxide is well known to be a mediator of inflammation and an inducer of apoptosis. Expression or activation of NADPH oxidase is increased in diabetes or high-glucose treated endothelial cells.13-15 We previously showed that long term treatment with the antioxidant N-acetylcysteine (NAC) could inhibit NADPH oxidase activation in the heart of diabetic rats, and it also abolished the compensatory increase in the antioxidant enzyme superoxide dismutase (SOD).16 It is, however, unknown whether or not short term NAC treatment may preserve the compensatory increase of endogenous SOD protein expression in diabetes, while maintaining its inhibitory effects on NADPH oxidase activation.
SOD, including cytosolic Cu/Zn-SOD, mitochondrial Mn-SOD and extracellular SOD,17 are enzymes involved in the scavenging of superoxide anions and play an important role in balancing ROS generation. Superoxide anions are mainly generated by the mitochondrial electron transport chain and are implicated in the progression of diabetes.18-20 Increased ROS production in diabetes can be prevented by the administration of SOD.21 A recent study showed that total SOD activity and extracellular Cu/Zn-SOD activity were reduced, whereas Mn-SOD activity was increased in the heart of long-term diabetic rats.22 This indicates that individual endogenous tissue antioxidant enzymes and potentially the total tissue antioxidant activity may change differently in diabetes. This complexity may potentially affect the effectiveness of antioxidant therapies. During early diabetes, various tissues may enhance their antioxidant enzymes activity to withstand hyperglycemia-induced ROS production. However, little is known regarding the tissue specific changes of overall antioxidant capacity and the underlying mechanism in early diabetes.
We hypothesized that changes in the levels of both NADPH oxidase and SOD may be tissue specific, possibly having a significant impact on the total tissue antioxidant activity and on the effectiveness of antioxidant treatment in early diabetes. Therefore, this hypothesis was tested in an established rat model of STZ-induced type 1 diabetes. Since glutathione (GSH) depletion may play an important role in the development of diabetic complications23 and GSH depletion occurs in early diabetes,24 we also explored the treatment effects of N-acetylcystein, a GSH precursor that can enhance the GSH antioxidant defense.25
MATERIALS AND METHODS
Experiment design and induction of diabetes
Sprague-Dawley male rats aged 8 weeks and weighing 250 to 280 gram were purchased from the Animal Unit of Wuhan University, China, and allowed to adapt in their cages for three days before experiments. All rats had free access to standard chow and water. The experiments were performed in accordance with our institutional animal care guidelines which are in line with the use of Laboratory Animals published by the US National Institutes of Health. The study was approved by Institutional Animal Care and Use Committee.
Diabetes was induced by a single tail vein injection (under halothane anaesthesia) of streptozotocin (Sigma, St. Louis, MO, USA) (STZ, 60 mg/kg body weight, freshly dissolved in 0.1 M citrate buffer, pH 4.5), while control rats (n=7) were injected equal volume citrate buffer alone. Three days following STZ injection, blood glucose levels were measured using a Glucose Analyzer (Beckman Instruments, Fullerton, CA, USA), and rats with hyperglycemia (set at plasma glucose ≥16.7 mM) were used for the experiments.
NAC, at the dose of 1.5 g/kg/day, had no effects on NADPH oxidase activity and SOD activity in non-diabetic control rats but had effects on these parameters in diabetic rats.16 In the current study, therefore, NAC was used only in diabetic rats. One week after the induction of diabetes, rats were randomly assigned into NAC-treated or untreated groups (n=7, per group), and they were treated respectively with the antioxidant NAC (1.5 g/kg/day, D+NAC) or an equal volume of 0.9% saline (D) delivered by oral gavage. Rats in the non-diabetic control group (C) were treated also with an equal volume of 0.9% saline delivered by oral gavage. Treatments in all groups were continued for four weeks.
Plasma and tissue preparations
At the end of four weeks of treatment, the rats were anti-coagulated with heparin (1000 IU/kg) and then anaesthetized with pentobarbital sodium (65 mg/kg body weight). Blood glucose level and body weight were measured by a Glucose Analyzer (Beckman Instruments, Fullerton, CA, USA) and a lab scale, respectively. Blood samples were collected in heparinized syringes by heart puncture after opening the thoracic cavity. The collected blood was immediately centrifuged at 3000×g, plasma separated, aliquoted into small microcentrifuge tubes, and stored at -80℃ for further detection. Heart, kidney, liver and lung tissues were harvested immediately after sacrifice and frozen in liquid nitrogen and stored at -80℃ for further experiments.
Inflammatory cytokines in plasma
Plasma levels of inflammatory cytokines tumor necrosis factor (TNF)-alpha and interleukin (IL)-6 were measured, using the commercially available rat ELISA kit (Bender Med, Vienna, Austria) according to the manufacturer's instructions.
Enzyme immunoassay (EIA) for free 15-F2t-isoprostane
15-F2t-isoprostane (15-F2t-IsoP), a specific marker of oxidative stress, was measured using an EIA kit (Cayman chemical, Ann Arbor, MI, USA). Thus, plasma samples or homogenized heart tissue (in PBS) were purified using Affinity Sorbent/Column (Cayman chemical, Ann Arbor, MI, USA) in the presence of 0.01% butylated hydroxytoluene and then processed for analysis of free 15-F2t-IsoP as described.26 The values of plasma or tissue 15-F2t-IsoP were expressed as pg/mL in plasma or pg/mg protein in tissue homogenates.
Total antioxidant activity determination
Tissue sections (50 mg) were homogenized in 1 mL of cold buffer (5 mM potassium phosphate, pH 7.4, containing 0.9% sodium chloride and 0.1% glucose) and were centrifuged at 10000×g for 15 min at 4℃. The supernatant was collected for assay. Total antioxidant activity in plasma and tissue homogenate was measured using the Total Antioxidant Status Assay Kit (Calbiochem, Darmstadt, Germany), which is dependent on the capacity in the sample to inhibit the oxidation of ABTS™ [2,2'-Azino-di-(3-ethylbenz-thiazoline sulphonate)] to ABTS™·+ by metmyoglobin (a peroxidase). The amount of ABTS™·+ produced can be monitored by reading absorbance at 600 nm. Under the reaction conditions used, the antioxidants in the sample suppress absorbance at 600 nm by the degree proportional to their concentration. Total antioxidant activity was calculated and expressed as millimole per liter (mM) in plasma or millimole per gram protein in tissue samples (mmol/g protein).
Total superoxide dismutase activity assay
SOD activity was determined using a Superoxide Dismutase Assay Kit (Cayman chemical, Ann Arbor, MI, USA) following the manufacturer's instructions. Plasma samples were assayed immediately after being thawed. Tissue sections (50 mg) were homogenized in 1 mL of cold 20 mM HEPES buffer, pH 7.2, containing 1 mM EGTA, 210 mM mannitol and 70 mM sucrose, and centrifuged at 1500×g for 5 min at 4℃, and the supernatants were collected for assay. The assay utilizes a tetrazolium salt for detection of superoxide radicals generated by xanthine oxidase and hapoxanthine. One unit of SOD is defined as the amount of enzyme needed to exhibit 50% dismutation of the superoxide radical. SOD activity was calculated and expressed as units per milliliter (U/mL) in plasma or units per milligram protein (U/mg protein) in tissue preparations.
Tissue sample preparations and western blot analysis
In order to characterize subcellular distribution of targeted proteins, tissue samples were processed to isolate membrane and cytosolic fractions as previously described.26 Briefly, 150 mg of heart, lung, kidney and liver tissue samples were pulverized separately with mortar and pestle in liquid nitrogen, and then homogenized using a Polytron homogenizer in 1.5 mL of cold buffer A, which contained (in mM) 50 Tris-HCl (pH 7.5), 5 MgC12, 10 EGTA, 2 EDTA, 1 NaHCO3, 1 PMSF, 1 β-glycerophosphate, 1 NaF, 1 Na3VO4, and (in µg/mL) 20 leupeptin and 4 aprotinin. The homogenate was centrifuged at 1000×g for 10 min at 4℃. The supernatant was collected and used as total tissue extracts. The supernatant was then further centrifuged at 100000×g for 60 min at 4℃. The supernatant was removed as cytosol fractions. The pellets were resuspended in buffer B containing 50 mM Tris-HCl (pH 7.5), 150 mM NaCl, 5 mM EDTA, 1% Triton X-100, 10% glycerol, 1 mM PMSF, 1 mM β-glycerophosphate, 60 mM Octyl β-glucopyranoside, 1 mM NaF, 0.1 mM Na3VO4, and (in µg/mL) 20 leupeptin and 4 aprotinin. The suspensions were homogenized again, held on ice for 30 min, and centrifuged at 100000×g for 60 min at 4℃. The supernatant was collected as membrane fractions. The proteins from each fraction (50-100 µg) were resolved via SDS-PAGE and subsequently transferred to PVDF membrane for immunoblot analysis. The membranes were blocked in 5% non fat milk for 2 h at room temperature, washed three times with TBS-T buffer each for 5 min, and then incubated overnight at 4℃ with primary antibodies raised against Cu/Zn-SOD (rabbit antiserum), Mn-SOD (rabbit polyclonal IgG) (1 : 1000, Millipore Corporation), p22phox and p67phox (rabbit polyclonal antibody, 1 : 500, Santa Cruz Biotechnology), respectively. After washing three times with TBS-T for 30 min, the blot was incubated for 1 h at room temperature with anti-rabbit IgG, HRP-linked antibody (1 : 10000, Cell Signaling). After washing the membranes, the blot was developed with enhanced chemiluminescence reagent (GE Healthcare) and then exposed to X-ray film. The same blot was stripped and reblotted with antibodies to ward GAPDH (1 : 2000, Cell Signaling) in total tissue extracts and cytosol fractions, or Na, K-ATPase (1 : 2000, Cell Signaling) in membrane fractions as an internal control. The intensity of the bands was quantified using image analysis software (Bio-Rad, Hercules, CA, USA), and the results were normalized to corresponding internal controls to correct for loading. Data are presented as percent change relative to the measurement in control rats.
Statistical analysis
The values are presented as mean±S.E.M. All biochemical assays were performed in duplicate and averaged before being analyzed. One-way or two-way analysis of variance was used for statistical analyses of data obtained within the same group and between groups, respectively. Tukey's test was used for multiple comparisons of group means. Significance was defined as p<0.05.
RESULTS
General characteristics and effects of NAC treatment
Administration of STZ resulted in characteristic symptoms of diabetes including hyperglycemia, and reduced body weight gain along with increased food and fluid intake when compared to age-matched controls (Table 1). Food consumption, water intake and plasma glucose levels in the diabetic group were much higher than those in the age-matched control group (p<0.01). NAC treatment significantly reduced water intake and food consumption compared with the diabetic untreated group (p<0.05), but did not significantly affect glucose levels. Body weight in the diabetic group was lower than that of the control group (p<0.05), and NAC treatment did not have significant effects on body weight gain (Table 1).
Oxidative stress marker: free 15-F2t-IsoP levels in plasma and tissue
Plasma (Fig. 1A) and tissue (Fig. 1B) levels of free 15-F2t-IsoP in the diabetic group were much higher than those in the control group (all p<0.05 or p<0.01). NAC treatment significantly reduced free 15-F2t-IsoP concentrations in the plasma and prevented the significant increase of 15-F2t-IsoP in the heart, lung and liver tissues seen in the diabetic group (all p<0.05, D+NAC vs. D). NAC significantly reduced, but did not normalize 15-F2t-IsoP levels in the kidney (p<0.05, D+NAC vs. D; p<0.05, D+NAC vs. C) (Fig. 1B).
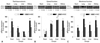
Total antioxidant activity in plasma and various tissues
Total antioxidant activity represents the total antioxidant capacity in tissues or in the plasma. Plasma total antioxidant activity in diabetic rats was higher than that of the control group (p<0.01) (Fig. 2A). NAC treatment significantly attenuated this increase in plasma total antioxidant levels in diabetic rats (p<0.05, D+NAC vs. D) (Fig. 2A). Changes in tissue total antioxidant activity and the effects of NAC treatment were tissue-specific. The total antioxidant activity in the heart in the diabetic group was higher than that in the control group (p<0.05) and was unaffected by NAC treatment (Fig. 2B). In contrast, however, total antioxidant activity in the lung, liver and kidney tissues was significantly decreased in the diabetic group as compared with control group (p<0.05) (Fig. 2B). NAC treatment elevated total antioxidant activity in the lung and liver (p<0.05, D+NAC vs. D), but not in the kidney.
Total SOD activity in plasma and various tissues
The amount of SOD present in the cellular and extracellular environments is crucial for prevention of ROS mediated oxidative cellular damage. Plasma total SOD activity in the diabetic rats was much lower than that in the control group (p<0.01) (Fig. 3A), NAC did not have significant effects on plasma total SOD activity. In the control rats, tissue total SOD activities in the heart and lung were much lower than those in the liver and kidney (Fig. 3B), suggesting that SOD might not be the predominant endogenous antioxidant enzyme in the heart and lung in rats under physiological conditions. In the heart, total SOD activity was slightly increased in diabetic rats as compared to control rats (p<0.05, D vs. C), and this compensatory increase in SOD activity was not significantly affected by NAC treatment. While total SOD activity in the lung, was slightly decreased in diabetic rats as compared to control rats, NAC prevented the reduction of SOD activity in the lung. Furthermore, total SOD activities both in the liver and kidney were decreased significantly in diabetic rats as compared to age-matched controls (all p<0.05) (Fig. 3B), and NAC treatment prevented the decrease in total SOD activity in the liver, but not in the kidney.
Protein expression of Cu/Zn-SOD and Mn-SOD
In the heart and lung tissues, the levels of Cu/Zn-SOD protein in the diabetic group increased significantly compared with controls (all p<0.05, D vs. C) (Fig. 4A). NAC attenuated this increase in Cu/Zn-SOD protein expression in the heart (p< 0.05, D+NAC vs. D), but did not significantly affect Cu/Zn-SOD protein expression in the lung (Fig. 4A). In contrast, however, Cu/Zn-SOD protein expression in liver and kidney tissues in diabetic rats was lower than that of age-matched control rats (p<0.05 in liver, p<0.01 in kidney) (Fig. 4A). NAC prevented the decrease of Cu/Zn-SOD protein in the liver and attenuated its reduction in the kidney (all p<0.05, D+NAC vs. D) (Fig. 4A).
The changes of Mn-SOD protein expression in various tissues were different from those of Cu/Zn-SOD. There was no significant difference in Mn-SOD protein expression in the heart tissue among control, diabetic and NAC-treated diabetic groups (Fig. 4B). However, Mn-SOD protein expression in both the lung and the kidney decreased significantly in the diabetic group as compared to the control group, and NAC prevented the reduction of Mn-SOD protein in the lung (p<0.05, D+NAC vs. D) but further decreased its level in the kidney (p<0.05, D+NAC vs. D) (Fig. 4B). It is of an interest to note that Mn-SOD protein expression in the liver increased significantly in diabetic rats as compared control rats, but this increase was not affected by NAC treatment (Fig. 4B).
Expression of NADPH oxidase subunit: p22phox and p67phox
NADPH oxidase has been shown to be a major source of ROS production,10,28 and its subunits p22phox and p67phox are increased in the myocardium of STZ-induced diabetic rats.16 In the present study, we detected p22phox and p67phox in various tissues. As shown in Fig. 5A, p22phox protein levels in the heart, lung and kidney of the diabetic group were significantly higher than those of the control group, and NAC prevented the increase of p22phox protein expression in the lung, but did not normalize their increase in the heart and kidney (all p<0.05, D+NAC vs. D) (Fig. 5A). While there was no significant difference in p22phox protein expression in the liver between control and diabetic rats, 4 weeks of NAC treatment decreased its expression significantly (p<0.05, D+NAC vs. D or C) (Fig. 5A).
The p67phox protein expression was significantly increased both in the cytosol and membrane fractions in the lung, liver and kidney tissues of diabetic rats relative to control rats (Fig. 5B and C), reflecting increased p67phox activation as well as increased total protein levels which were in line with the increases of total p67phox protein expression in these tissues (data not shown). However, p67phox protein expression in the heart was significantly increased in the membrane fractions while markedly decreased in the cytosol fractions, reflecting increased p67phox activation. NAC prevented the increase of p67phox membrane translocation in the heart and liver and significantly attenuated the increase of p67phox membrane translocation in the lung and kidney tissues (all p<0.05, D+NAC vs. D) (Fig. 5C).
DISCUSSION
We have shown in the current study that the level of 15-F2t-IsoP, a biomarker of oxidative stress,29,30 was elevated both in plasma and various tissues of type 1 diabetic rats. In contrast to reductions in total antioxidant activity in the lung, liver and kidney tissues, total antioxidant activity in the heart was elevated in the diabetic rats accompanied by an increase in heart tissue total SOD activity which indicated a significant up-regulation of self-defense mechanisms in the heart. The variations in SOD activity and protein expression in diabetes were tissue-specific. NAC treatment up-regulated Mn-SOD protein expression in diabetic rats in the lung tissue, but unexpectedly reduced Mn-SOD protein expression in the kidney. The underlying mechanisms governing the tissue-specific effects of NAC on SOD protein expression are not clear, nevertheless, it might be due to tissue-specific effectiveness of NAC in reducing the production of superoxide.16 Consequently, the organism may regulate the levels of SOD activation or protein expression on a basis of needs. It should be noted that NAC uniformly reduced the increase of p22phox protein expression and p67phox activation in all tissues studied, most likely inhibiting the activation of NADPH oxidase, a major source of ROS production in diabetes. This result is keeping with findings of a previous study which showed that NAC can inhibit NADPH oxidase activity in osteoblast precursor cells.31 Thus, our present studies together with others suggest that NAC can break vicious cycle of ROS-induced ROS production via NADPH oxidase activation in diabetes.
Increased oxidative stress can be the result of several diabetes-induced abnormalities, including auto-oxidation of glucose, the formation of advanced glycation end products, and impairment of defense system.32 The impaired defense system is always associated with reduced antioxidant capacity, abnormal activity or expression of antioxidant enzymes in diabetes.33-35 Among various antioxidant enzymes, SOD plays an important role in balancing ROS generation and the overall tissue antioxidant capacity. In our study, total antioxidant activity and SOD activity in the lung, liver and kidney tissues and total SOD activity in plasma were compromised in diabetic rats. It should be noted total antioxidant activity both in the plasma and heart tissues and total SOD activity in heart tissues were increased in diabetic rats, reflecting increased systemic antioxidant capacity overall and in the heart in particular. However, heart tissue level of 15-F2t-isoprostane was increased despite these increases in systemic and heart tissue antioxidant activity. This indicates that up-regulation of antioxidant defenses is not sufficient to withstand hyperglycemia-induced increase in ROS production. It is also worth to note that in the control rats, the tissue total SOD activities in the heart and lung were much lower than those in the liver and kidney, which may suggest that SOD might not be the predominant endogenous antioxidant enzyme in the heart and lung in rats under physiological conditions. Other endogenous antioxidant enzymes such as glutathione peroxidase also play important roles in maintaining tissue antioxidant activity in these tissues under physiological conditions as well as in diabetes.36 Indeed, glutathione peroxidase and catalase are increased in the heart and lung of diabetic rats.36 This may explain the discrepancy between total antioxidant activity and SOD activity in plasma observed in the current study.
NADPH oxidase, a transmembrane enzyme located in intracellular organelles, is one of the major sources of ROS in diabetes.9,10 Recent studies show that the NADPH oxidase p22phox and gp91phox subunits were over-expressed in rat aorta,37 and that the p22phox, p47phox and p67phox subunits were increased in testes in STZ-induced diabetic rats.28 Our recent study shows that p22phox and p67phox, but not the p47phox, are robustly enhanced in the diabetic myocardium, and that NAC can prevent their increase.16 Therefore, in the current study, we specifically examined the changes of p22phox and p67phox in various tissues in diabetic rats, and found that p22phox and p67phox subunits were over-expressed or activated in various tissues in early diabetic rats, and that NAC treatment prevented or attenuated these changes in diabetes. NAC mediated reduction of p67phox translocation to membrane in various tissues was seen in the current study. This finding is in keeping with a previous study showing that NAC can inhibit NADPH oxidase activation.10 Thus, inhibition of NADPH oxidase over-expression and activation and the subsequent ROS production may represent a mechanism by which NAC reduces oxidative stress in diabetes in addition to its ROS scavenging property.
Mitochondrial dysfunction is another major intracellular source of ROS through the mitochondrial respiratory chain (MRC).10 About 4% of oxygen consumed in the mitochondria is converted to ROS.38 Impaired activities of enzymes in the MRC and mitochondrial DNA mutations were detected in patients with diabetes and in animal models of diabetes.39 Mitochondrial derived superoxide anion is viewed as a single unifying mechanism for diabetic complications.18,40 Over-expression of Mn-SOD, the major scavenger of mitochondrial superoxide anion, prevents high glucose-induced oxidative stress, cell apoptosis and mitochondrial DNA damage in retinal endothelial cells,19,20 and protects the retina from diabetes-induced oxidative stress in mice.41 In our present study, Mn-SOD expression in diabetic lung and kidney tissues decreased significantly, whereas Mn-SOD expression in diabetic liver tissue increased but was unchanged in heart tissue. This suggests that the change in mitochondrial SOD in response to hyperglycemia-induced oxidative stress is tissue-specific in early diabetes. NAC increased Mn-SOD in the lung, however, it did not increase it in the liver and further reduced its protein expression in the kidney, which might be the reason of why NAC could not bring down kidney tissue 15-F2t-IsoP content to a level comparable to that in the control rats.
Inflammation has been widely recognized as a key process of the progression of diabetes.42,43 Some inflammatory factors, such as TNF-alpha and IL-6, are elevated in patients with type 2 diabetes,44 and correlate with the incidence of diabetic macrovascular complications, especially atherosclerosis development.45 Our previous studies showed that long term (8 weeks) treatment with NAC attenuated myocardial levels of IL-6 protein expression and reduced plasma level of TNF-alpha in STZ induced diabetic rats.16,46 In the present study, we found that plasma levels of TNF-alpha and IL-6 were significantly increased in STZ-induced diabetic rats early at 4 weeks after the establishment of diabetes, and that NAC treatment for 4 weeks decreased the secretion of TNF-alpha and IL-6 cytokines, which reflects a suppression of inflammatory responses. Similar to our study, NAC has been demonstrated to have anti-inflammatory property both in vitro in lipopolysaccharide-activated macrophages47 and in vivo in humans.48
In summary, the current study shows that oxidative stress occurs at an early stage of diabetes, and changes in the major pro-oxidant enzyme NADPH oxidase and the antioxidant enzyme SOD are tissue-specific in STZ-induced type 1 diabetic rats. Antioxidant NAC can confer protection against oxidative stress by suppressing NADPH oxidase activation and restoring or enhancing SOD enzyme activity and/or protein expression. The effects of NAC are tissue-specific, being more effective in the heart, lung and liver and less effective in the kidney. The current findings may have potential clinical implications in developing therapies for the treatment of diabetic complications.


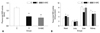



 XML Download
XML Download