

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
The p16 gene is one of the cell cycle regulating genes and encodes a nuclear protein, p16 which inhibits the D-type cyclin/cyclin-dependent kinase complexes that phosphorylate the retinoblastoma gene product (pRb), thus blocking G1-S cycle progression (1, 2). The inactivation of p16 tumor suppressor gene promotes cell proliferation, and is found in many different types of carcinomas such as gastric carcinoma, bladder tumor, glioma, breast cancer and head and neck tumors (3). There is compelling evidence that the inactivation of p16 is an important genetic event in immortalization of keratinocytes (4). In previous studies of the p16 in cervical carcinomas, methylation specific polymerase chain reaction (PCR) has shown a high level of methylation (1, 5), concordant with reports that the p16 gene is frequently inactivated through methylation rather than mutation or deletion. DNA methylation is a frequent epigenetic event in many human cancers (6, 7), and the factors inducing methylation remain unclear, but structural abnormalities of local DNA, and exposure to heavy metals are only known causes (8). A growing number of cancer-related genes are being recognized for the presence of dense methylation of cytosine in normally unmethylated CpG-rich sequences, called CpG islands within the 5' gene promoter regions.
Besides p16 gene inactivation, viral infection also participates in the dysregulation of cell cycle. A clear relationship between human papillomavirus (HPV) and cervical squamous cell carcinomas (SCCs) is well established; HPV-infected invasive SCCs tend to express more immunoreactivity for p16 protein or p16 methylation status than do HPV-negative cervical carcinomas, which is now regarded as surrogate biomarkers of HPV infection along with Ki-67 labeling index (5, 9, 10). Epstein-Barr virus (EBV) was suggested as another oncogenic virus in cervical carcinogenesis, based on findings of clonal nature of EBV in cervical carcinoma cells and the presence of EBV in precancerous lesions of the cervix (11). Since then, much debate has been evoked because of divergent results including a relatively high prevalence rate of EBV-positive nonneoplastic cervical tissue, which shed doubt on the possibility. However, heterogeneous reports on lymphoepithelioma-like carcinoma of the uterine cervix in Asian women showing a higher EBV infection (12), and the co-work of HPV and EBV cannot exclude EBV's direct, or indirect impact on cervical carcinogenesis (7, 11, 13). Considering that the tumors like gastric or nasopharyngeal carcinomas, which are known to be closely related with EBV's oncogenic effects, were identified in a high frequency of p16 methylation (14, 15), a close relationship exists between EBV and p16 gene. In Korea, one study about HPB and p16 methylation has been retrieved (16), and few studies about EBV detection or p16 alterations in cervical carcinomas have been retrieved (17, 18). There is, however, virtually no published information on the methylation changes during the multistage pathogenesis of EBV-related cervical lesions.
In this study, we explored EBV infection of the uterine cervix with reference to p16 gene that is frequently methylated in cervical carcinomas. Moreover, we investigated as to whether the two factors play an independent or synergic role during cervical carcinogenesis.
MATERIALS AND METHODS
Tumor samples
Eighty-two, formalin-fixed, paraffin-embedded cervical lesions were used for immunohistochemical, polymerase chain reaction (PCR) and in situ hybridization (ISH) studies. Materials were obtained from the pathology files of Anam Hospital, Korea University College of Medicine between 1995 and 2001. Samples included 41 cases of SCCs, 30 cases of cervical intraepithelial neoplasms (CIN 1; 11 cases, CIN II; 3 cases, CIN III; 16 cases), and 11 cases of non-neoplastic cervices.
DNA extraction
DNA samples were extracted from several serial six µm-thick paraffin sections as previously described (19). Briefly, tissue samples were treated with lysis buffer containing 100 µg/mL proteinase K (Merck, Darmstadt, Germany) at 48℃ for 48 hr. DNA extraction was then performed by phenol/chloroform treatment and precipitation with ethanol. The quality of the DNA extracted was confirmed with a beta-globin gene-specific primer pair (BGLO1 and BGLO2), which amplifies 110 base pairs.
Bisulfite modification and methylation-specific polymerase chain reaction (MSPCR) for detecting p16
DNA methylation patterns in the CpG islands of the p16 were determined by MSPCR assays by Herman et al. (20). Briefly, 200 to 300 ng DNA was first treated with 3 mol/L sodium bisulfite to convert nonmethylated cytosine to uracil residues. As a diagnostic step, we used heminested PCR to increase the sensitivity of detection with primers p16M/p16 mol/L2r for the first and p16Mf/p16Mr for the second step under conditions as described. Base sequences of used primers are as follows;
p16-methylated (p16-M):
p16-unmethylated (p16-U):
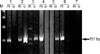
In all cases, the amplification of nonmethyl-specific alleles served as controls for the efficiency of cytosine conversion and DNA quality. Controls without DNA and positive controls for U and M reactions were performed for each set of PCRs. The PCR product was visualized in agarose gels stained with ethidium bromide under ultraviolet illumination. DNA methylation was determined by the presence of a150-bp and 151-bp fragments in those samples amplified with the p16-M and p164-U primers, respectively. Positive PCR products in p16-M and/or p16-U are interpreted as methylated, and PCR products in only p16-U are interpreted as unmethylated status.
Immunohistochemistry for p16
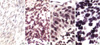
Immunohistochemical staining of p16 was performed with p16 monoclonal antibody (SC 468; Santa Cruz Biochemicals, CA, U.S.A.) diluted at 1:50. Immunodetection was performed with biotinylated antimouse immunoglobulins, followed by peroxidase-labeled streptavidin, LSAB-2 (DAKO, Glostrup, Denmark) with diaminobenzidine chromogen as the substrate. Each section was counterstained with hematoxylin. Incubation omitting the specific antibody, as well as with unrelated antibodies, was used as a control of the technique. A positive control was included with each slide. The results were interpreted as positive according to the established criteria by Geradts et al. (21).
PCR for detection of EBV
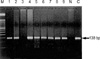
To examine the presence of EBV DNA in the samples, 0.1 µg DNA from a cervical sample was subjected to PCR analysis using EBNA-1 primers which target a 138 base pair segment of the Bam H1W internal repeat (IR 1) region described by Coates et al. (22). Base sequences of primers were as follows;
Thirty-five PCR cycles at 95℃ for 7 min, at 94℃ for 1
min, at 58℃ for 40 sec, and at 72℃ for 40 sec were carried
out. And it was carried out at 72℃ for 7 min Polymerase
chain reaction was carried out in duplicate or triplicate. The amplified products were run on 2% Nuieve/agarose gel and were transferred onto a Hybond N+ membrane (Amersham Biosciences, Uppsala, Sweden). They were stained with ethidium bromide and examined under ultraviolet illumination. As a positive control of EBNA-1, B95-8 cell line was used.
In situ hybridization (ISH) for EBV
EBV ISH was performed using oligonucleotide probes against EBV-encoded RNA-1 (EBER-1; Novocastra, Newcastle, U.K.), following previously reported procedures (23). EBER-1 positivity was shown by latently EBV-infected cells. Briefly, EBER-1 ISH was performed on each case using digoxigenin-labeled riboprobes produced from plasmid templates. A step-by-step description of one of the testing protocols, which uses EBER-1 was executed on each case and hybridized overnight. RNase was used to facilitate the washing away of unbound probe and a detection system based on the application of antibody to digoxigenin was linked to alkaline phosphatase. It was counterstained with hematoxylin. Known cases of EBV-harboring tonsils were used as controls. Strong signals of the nuclei were interpreted as positive results.
Data analysis
We divided the cases into four groups as follows: non-neoplastic normal cervical tissue, low grade dysplastic lesions (cervical intraepithelial neoplasm, CIN I), high grade lesions (CIN II and III) and invasive SCCs. Subsequently, we compared EBV infection with p16 methylation and p16 protein expression. Statistical analysis was performed by means of SPSS software version 10. The associations between the discrete variables were assessed using McNemar's exact test and chi-square test. The same tests were used to evaluate the influence of the tumor grade of CIN. Differences were considered statistically significant for p<0.05.
RESULTS
All the results of p16 methylation, p16 protein expression, EBNA-1 PCR and EBER-1 ISH of the eighty-two cases were shown in Table 1.
p16 methylation status
Non-neoplastic cervices showed unmethylation in all the cases, but 40% (12/30) of cervical intraepithelial neoplasms and 61% of invasive squamous cell carcinomas (25/41) showed p16 methylation (p=0.003, Fig. 1, Table 1). There existed statistical differences among CIN I, II and III (p=0.03, data not shown), while the advanced stage of squamous cell carcinomas (stage IIb) revealed a higher p16 methylation rate than the early stage (I, IIa) of squamous cell carcinomas, however, the results were not statistically significant (p=0.107, Table 2).
Immunohistochemistry for p16 protein
Non-neoplastic cervices were immunonegative for p16 protein except for basal cells that are known to normally express p16 protein, but 53% of cervical intraepithelial neoplasm (16/30 cases) and 68.3% of invasive squamous cell carcinomas (28/41) expressed p16 protein. These results were significantly different among the four groups (p=0.001, Table 1, Fig. 2). However, no differences existed among the three grades of cervical intraepithelial neoplasms (p=0.600, data not shown). In comparison of invasive carcinomas according to the stage, there was a higher expression rate in more advanced stages, but no statistically significant differences existed (p=0.192, Table 2).
EBV detection
EBV was detected by EBNA-1 PCR in 9.1% of non-neoplastic cervical tissue (1/11), 36.7% of cervical intraepithelial neoplasm (11/30) and 36.6% of invasive squamous cell carcinomas (15/41) (Fig. 3). There was no statistically significant difference in the detection rate of EBV among the four groups (p=0.331, Table 1), whereas a significant difference existed between the two groups (non-neoplastic cervices vs. precancerous CIN and cervical cancers, p=0.040, data not shown). There were no statistical differences among the grades of CIN (p=0.747, data not shown).
Exact test of concordance of p16 MSPCR with the immunohistochemistry of p16 protein, and the EBV detection rate
By McNemar's exact test, p16 MSPCR and the immunohistochemical results were proven to be concordant, while the detection rate of EBV was different between in situ hybridization and PCR.
Correlation among p16 methylation, p16 immunoreactivity and EBV detection
The cervical lesions positive for EBV by PCR showed a higher rate of p16 methylation and/or p16 immunopositivity, which was statistically significant (Table 3).
DISCUSSION
In this study, 61% of squamous cell carcinomas of the uterine cervix showed p16 methylation, which was higher than 31% in the report of Wong and his colleagues (2). Cervical carcinomas are known to show a high proportion of p16 methylation, while other gynecological malignancies have variable frequencies (24). Sano et al. (4) reported that invasive squamous cell carcinoma and CIN lesions (low and high grades) exhibited higher p16 immunoreactivity than non-neoplastic cervices. Based on the results that dysplastic lesions progressed toward carcinomas with increasing p16 methylation, methylation has been thought to play a critical role in oncogenesis as in lymphoma or carcinomas of other organs (25). However, the frequency of p16 methylation between early and advanced stages showed no statistically significant difference, which is quite different from those of the tumors in the head and neck, breast or bladder (2, 26, 27). This result may indicate that early cervical carcinogenesis requires p16 methylation, but additional molecular genetic events are necessary for its progression.
Variable data on p16 immunoreactivities and cervical carcinomas have been described in the literature; most of them reported loss of p16 protein (28), while some reported the reexpression of p16 protein (29). In our study, dysplastic cervical epithelium and squamous cell carcinomas showed a higher expression of p16 protein in the cervical epithelium, which contradicts most of the previous studies, but fits well with that of Klaes et al. (29). The statistically concordance despite discrepancy in individual comparison between p16 immunohistochemistry and p16 MSPCR in this study indicates that the former may employ an inexpensive screening method to detect squamous cells showing early dysplastic changes. Interpretation of immunoreactivity for p16 protein has some limitations because p16 expression is variable in normal, dysplastic epithelium and neoplastic tissue; basal layer of nonneoplastic exocervix expresses weak and focal nuclear stainability (30). Theoretically, p16 protein should be expressed only in the nuclei but cytoplasmic staining pattern was occasionally observed, and its significance remains unclear. Cytoplasmic immunoreactivity may be induced by a nonspecific uptake or may be caused by functional heterogeneity (21). Some of the cultured cells pertaining to p16 gene show nuclear negativity but with variable cytoplasmic reactivities, or cytoplasmic p16 detected by immunoblotting may indicate that this cytoplasmic localization of p16 is "dormant" like that of the tumor suppressor gene, which has functions different from the nuclear one (31). A discrepancy in the results between the two detection methods for p16 inactivation was noted in our series. MSPCR has been known to be more sensitive and specific than Southern hybridization (20). In contrast, immunohistochemistry using p16 monoclonal antibody is simple and useful but has several technical limitations. For example, p16 protein is prone to break during the processing, and heterogeneous immunoreactivity in nonneoplastic tissue could cause confusion during interpretation. Progressive increase of p16 immunoreactivity and methylation were observed at carcinomatous progression. Timmermann et al. (32) demonstrated that epithelial cells gained increased activity of beta-galactosidase that is associated with senescence and produced p16 protein with no normal suppressor function. The reexpression of this incomplete protein detected by immnohistochemical method may be explained (33).
In this study, 60.5% of EBV-positive cervical tissues were p16-methylated, whereas 68.2% of EBV-negative cervical tissues were p16-unmethylated. These results were similar to those of nasopharyngeal undifferentiated carcinoma or EBV-positive gastric carcinomas, where there was likewise a trend toward an association between EBV infection and loss of p16 expression. One possible explanation for the association of p16 loss and EBV infection is that EBV-infected tumors could have a higher growth rate and therefore a greater opportunity to accumulate more mutations, including those of p16. In this study, we confirmed EBV infection by PCR and ISH. There was disconcordance between the results through EBV PCR and ISH. In general, the amplification of DNA through PCR was possible even in one viral particle, thus, theoretically it is a more sensitive method than ISH (9). In practice, a sensitive and specific practical method for detecting EBV is ISH (34). Cases of EBV-negativity by PCR but EBV-positive reaction by ISH in this study, seemed to be caused by insufficient amplification process. In this study, 39% of SCCs were EBV-positive, which was not a high frequency compared to non-neoplastic tissues or CIN. However, EBV-positive carcinomas showing a high rate of p16 methylation suggested that EBV alone does not play a key role in uterine cervical squamous epithelium, rather it may be involved in the earliest step of oncogenesis, which is somewhat dissimilar to that of nasopharyngeal carcinoma or Burkitt's lymphoma. Recent development of molecular genetics for carcinogenesis and virus, methylation of CpG islands possessing p16 is induced by the integration of viral DNA into host cells (34, 35). It was suggested that methylation of foreign DNA integrated with host DNA or adjacent host DNA, especially cell cycle regulatory site is the first step of carcinogenesis (36), which is similar to those of SV40 in relation to malignant mesothelioma or EBV in relation to EBV-positive gastric carcinomas (15, 37). Methylation of EBV genome inhibits suppression of immunodominant viral antigen by EBV-infected tumor cells and escapes immunosurveillance (15). Pharmacological blocking by methylation inhibitor causes tumor suppressor gene to be protected from aberrant methylation, might finally gain its own function, which the tumor cells expose immunosurveillance system. Actually, clinical application is now being attempted (38). On this mechanism, the relationship between EBV infection and p16 methylation plays a role in the early change of cervical carcinogenesis, and may provide promise for future treatments.
In conclusion, we did not find an association between p16 methylation status and stage. These observations well corresponded to the fact that p16 inactivation is an early epigenetic event in cervical cancer. A more intense reactivity of p16 protein was associated with p16 methylated status in this study, which might imply that p16 immunoreactivity represents the reactivation of senescent p16 protein in cancers, keeping with a previous report of cervical carcinoma, but differs from the correlation with the advanced stage of cervical SCC in the earliest study of Wong et al. (2). Although our findings need to be extended to a larger series, the pattern of p16 methylation in cervical lesions with or without EBV-infection may help identify subgroups at increased risk for accelerated progression.




 XML Download
XML Download