

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Nonalcoholic fatty liver disease (NAFLD) is rapidly becoming one of the most prevalent liver diseases, accounting for 25.24% of the general population worldwide [1]. The theory of the developmental origins of health and disease has linked the changes in early life nutrition to the susceptibility to metabolic disorders in the offspring [23]. The maternal obesity increased the risk of biopsy-proven NAFLD in the offspring by 2.26-fold [4]. The neonatal offspring of obese mothers with gestational diabetes mellitus had a 68% increase in the lipid content in the liver measured by magnetic resonance imaging [5]. These findings underscore that nutrition during pregnancy and lactation are critical windows for the susceptibility to NAFLD in offspring. The pathogenesis of NAFLD remains unclear. Hence, there is a lack of nutritional interventions to prevent and treat NAFLD.
NAFLD consists of a spectrum ranging from simple steatosis (SS) and steatohepatitis to fibrosis, cirrhosis, and eventually hepatic carcinoma. Nitric oxide dioxygenase-like receptor family pyrin domain containing 3 (NLRP3) inflammasome may play a critical role in the progression of NAFLD [6]. NLRP3 inflammasome is a multicomplex protein composed of NLRP3, apoptosis-associated speck-like protein containing C-terminal caspase recruitment domain (ASC), and pro-Caspase-1. NLRP3 inflammasome produces active Caspase-1 and releases inflammatory cytokines, including interleukin (IL)-1β and IL-18, mediating the inflammatory response of the liver. NLRP3 activation promotes the development of NAFLD [7]. A high-fat diet (HFD) or a combination with streptozotocin (STZ) in rodents stimulated the activation of NLRP3 inflammasome and mediated the secretion of IL-1β and IL-18 [89]. Hence, it was speculated that the activation of NLRP3 may occur in the livers of offspring mice from HFD and STZ-administered dams, which has not been reported before.
Betaine is a natural compound abundant in many foods, such as quinoa, oats, spinach, and beets, and can be consumed directly through the diet. Our previous studies have shown that betaine supplementation could relieve lipid accumulation and alleviate steatosis in mice induced by a HFD [10] and apolipoprotein E knockout [11]. We further investigated that maternal betaine intake alleviated hepatic steatosis in the offspring induced by maternal HFD [12]. The anti-inflammatory effects of betaine are closely related to the inhibition of NLRP3 inflammasome [13]. The betaine treatment inhibited the activation of the hepatic NLRP3 inflammasome pathway by suppressing the Caspase-1-dependent maturation of IL-1β and IL-18 and IL-6 production. It contributed to the alleviation of liver injury in fructose-fed rats [14]. On the other hand, it is unclear if maternal betaine supplementation alleviates the fatty liver in the offspring through regulations on NLRP3 inflammasome.
Low STZ doses damage pancreatic β-cells, inducing hyperglycemia and diabetes [15]. Combined with a HFD, this method can establish a model for nonalcoholic steatohepatitis (NASH) [161718]. Therefore, a HFD and STZ during pregnancy and lactation were used to investigate the lipotropic and anti-inflammatory effects of maternal betaine supplementation in the livers of the offspring mice. In addition, this study further investigated whether the underlying mechanism is related to regulations on hepatic NLRP3 inflammasome. This study contributes to a dietary strategy for the management of fetal-derived NAFLD.
MATERIALS AND METHODS
Animals experiment
Four-week-old female C57BL/6J mice were purchased from the experimental animal company of Shandong Jinan Pengyue (SCXK 2014-0007). The mice were fed a standard diet (MD12031) during a one-week adaptation period. The mice were assigned randomly to 3 groups and subjected to different feeding methods: control diet (CD), fatty liver disease (FLD), and fatty liver disease + 1% betaine (FLD-BET). The CD group was maintained on the standard diet. The mice in the FLD and FLD-BET groups were switched to a HFD (45 kcal% fat, MD12032). Supplementary Table 1 lists the composition of the standard diet and HFD. After 3 weeks, all the mice were mated to achieve pregnancy. The pregnant mice in the FLD and FLD-BET groups were injected subcutaneously with 55 mg/kg STZ once daily for 3 consecutive days. The fasting blood glucose (FBG) concentrations were measured 8 days later, and mice with FBG concentrations higher than 6.1 mmol/L were diagnosed with hyperglycemia and selected for further study. The mice in the CD group were given an equal volume of citrate buffer. Betaine at 1% wt./vol. was added to the drinking water (tap water) provided to the FLD-BET group after pregnancy. All pregnant mice were fed the indicated diet until the offspring mice were weaned. The offspring mice were breast-fed for 3 weeks and then given a standard diet for 5 weeks. Both the standard diet (MD12031) and HFD (MD12032) were purchased in Medicience (Jiangsu, China). Betaine was obtained from Danisco A/S (Copenhagen, Denmark). All experimental mice were housed in the experimental animal center of Jinan University under a temperature- and humidity-controlled environment. The mice were raised on laminar flow shelves in cages and provided food and water ad libitum. The fasting weights and food and water consumption were measured weekly. The cages and drinking bottles were washed and disinfected weekly. At the end of the experiment, all the mice were fasted for 12 h and sacrificed after anesthesia. The livers and blood samples were collected and stored at −80°C. Six to ten mice samples per group were chosen for subsequent experiments. Fig. 1 shows the design of the animal experiment.
Serum biochemical parameters
The serum biochemical parameters were determined using an automatic biochemical analyzer. The measured parameters included alanine aminotransferase (ALT), aspartate transaminase (AST), glucose (GLU), high-density lipoprotein cholesterol (HDL-C), low-density lipoprotein cholesterol (LDL-C), total cholesterol (TC), and triglyceride (TG).
Measurement of NAFLD
The liver TG content was measured using a commercially available kit (E1003-2; Applygen Technologies Inc., Beijing, China). For histology analysis, the liver samples from each mouse were divided into several sections. One sample per mouse was fixed in 10% formalin and embedded in paraffin. The sections were sliced from the paraffin blocks and stained with hematoxylin and eosin (HE) to evaluate the pathology of the hepatocytes. Another section was embedded directly in the optimal cutting temperature compound, sliced, and stained with oil red O to examine the lipid droplets. The degree of hepatic steatosis was quantified according to the Nonalcoholic Steatohepatitis Clinical Research Network Scoring System. The steatosis severity was divided into 4 grades based on the proportion of steatosis to the entire lobule (grade 0: < 5%; grade 1: 5–33%; grade 2: > 33–66%; grade 3: > 66%). Inflammation grading was classified into 3 levels based on the number of inflammatory cell piles (grade 0: 0; grade 1: < 2; grade 2: 2–4; grade 3: > 4) [19].
Determination of one carbon metabolite concentrations in the liver
The concentrations of choline, betaine, homocysteine (Hcy), trimethylamine-n-oxide, s-adenosine methionine (SAM), and s-adenosine homocysteine (SAH) in the livers were detected by high-performance liquid chromatography, as described in previous work [10].
RNA isolation and real-time polymerase chain reaction (RT-PCR) analysis
The total RNA was isolated from liver tissue using a total RNA extraction kit (CW0581S; Cwbio, Beijing, China). Gene expression analysis was conducted quantitatively by RT-PCR on an Applied Biosystems 7500 rapid RT-PCR system (Applied Biosystems, Foster City, CA, USA). The data were analyzed using the method of 2−ΔΔCT and are expressed as the fold difference of the gene of interest relative to β-actin. Supplementary Table 2 lists the primers used to detect the target genes and β-actin.
Western blotting analysis
The proteins were extracted from the liver samples using a total protein extraction kit (CW0891M; Cwbio). The protein concentrations were quantified using a bicinchoninic acid assay kit (P0012; Beyotime, Shanghai, China). The sample proteins (50 μg) were separated by 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis and transferred to polyvinylidene difluoride membranes (IPVH00010; Millipore, Burlington, MA, USA). Subsequently, the membrane was blocked with a blocking solution and probed by incubation with primary antibodies for NLRP3 (15101; Cell Signaling Technology [CST], Danvers, MA, USA); ASC (67824; CST); Caspase-1 (24232; CST); Cleaved Caspase-1 (89332; CST); Anti-IL-1β (ab234437; Abcam, Waltham, MA, USA); Anti-IL-18 (ab207323; Abcam); GAPDH (ab181602; Abcam) at 4°C overnight. The membrane was washed 3 times with Tris-Buffered Saline and Tween 20 and incubated with horseradish peroxidase-conjugated secondary antibodies for 1 h, followed by detection using enhanced chemiluminescence (WBKLS0100; Millipore).
DNA isolation and determination of global DNA methylation
Liver samples were extracted to obtain high-quality genomic DNA using a TIANamp Genomic DNA kit (DP304; TIANGEN, Beijing, China). Global DNA methylation was measured using a MethylFlash Global DNA Methylation (5-mC) ELISA Easy Kit (P1030; EpiGentek, Farmingdale, NY, USA) according to the manufacturer’s instructions.
Measurement of DNA methylation levels of gene promoters
DNA samples were modified by bisulfite using the ZYMO EZ DNA Methylation-Gold Kit (D5006; ZYMO RESEARCH, Irvine, CA, USA). The gene promoter sequence of mice was obtained from the website: http://genome.ucsc.edu/cgi-bin/hgNear. The bisulfite genomic sequencing PCR amplification primer of each gene was designed according to the website: http://www.urogene.org/cgi-bin/methprimer/methprimer.cgi; Supplementary Table 2 lists the sequences. The modified DNA samples were amplified by PCR. For each 50 μL PCR reaction, 5 μL of 10× buffer, 4 μL of dNTP, 0.25 μL of Taq Hotstart polymerase (Takara, San Jose, CA, USA), 2 μL of 10 μM forward primer, 2 μL of 10 μM reverse primer, 4 μL of DNA template, and 32.75 μL of water were used. The temperatures program was as follows: 95°C for 3 min followed by 40 cycles of 95°C for 30 s, Tm for 30 s, 72°C for 30 s, and 72°C for 10 min. Take 10 μL amplified products for gel electrophoresis. The amplification products were sequenced using the dideoxy chain termination method, and the methylation level was determined.
Statistic analysis
SPSS 24.0 (IBM Corp., Armonk, NY, USA) was used for data analysis, and GraphPad Prism 7.0 (GraphPad Software Inc., San Diego, CA, USA) was used to perform graphs. All normally distributed quantitative data were represented as the means ± SEs of the means. Multiple comparisons between the 3 groups were performed using a one-way analysis of variance and the Bonferroni test. The relationships between the contents of one carbon metabolite and the expressions of NLRP3 inflammasome were evaluated by linear correlation analysis. A P-value of < 0.05 was considered significant.
RESULTS
Effects of maternal betaine supplementation on the visceral weight and biochemical characteristics in mice
The dietary intakes in the dams and offspring were similar in the 3 groups (Supplementary Fig. 1). The litter size was also similar, with an average of 7–10 per litter. In dams, the FLD group had significantly higher final body weights (P = 0.003), body weight gains (P = 0.001), and body weight gain/initial body weight ratio (P = 0.001) than the CD group (Supplementary Table 3). The weight of the offspring mice increased with time, while no significant difference existed among groups (Fig. 2). On the other hand, the liver/body ratio (P = 0.017) and visceral fat/body ratio (P = 0.005) were lower in the FLD-BET group than in the FLD group (Table 1). The serum ALT level in the offspring mice was significantly higher in the FLD group than in the CD group (P < 0.001). Maternal betaine intervention significantly reversed this trend (P < 0.001). By contrast, maternal betaine supplementation did not affect the serum levels of the other parameters, including AST, TG, TC, LDL-C, HDL-C, and GLU in offspring mice (Table 1).
Fig. 2
Body weight of offspring mice. Data are presented as the means ± SEs of the means.
CD, control diet; FLD, fatty liver disease; FLD-BET, fatty liver disease + 1% betaine.
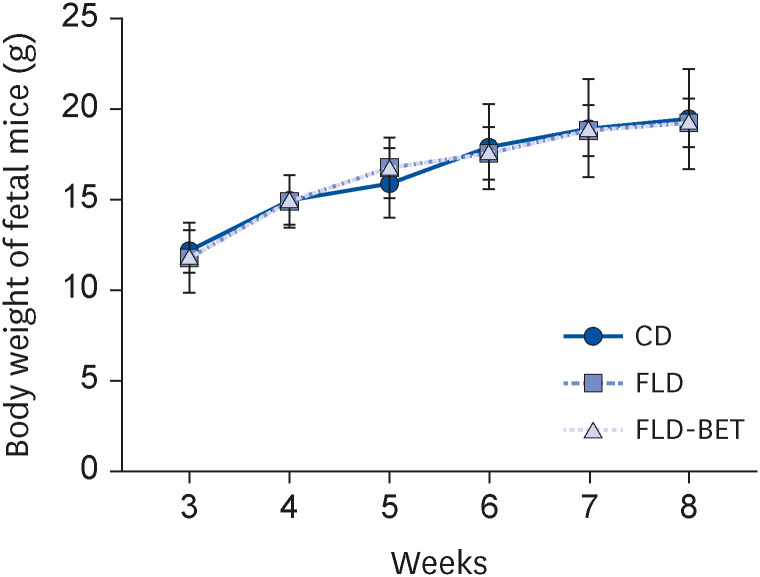
Table 1
Effects of maternal betaine supplementation on the visceral weight ratio and serum biochemical parameters of offspring mice
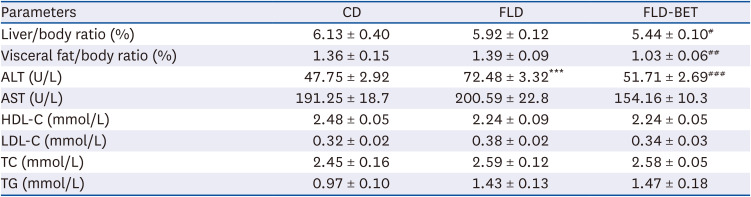
Data are presented as the means ± SEs of the means.
CD, control diet; FLD, fatty liver disease; FLD-BET, fatty liver disease + 1% betaine; ALT, alanine aminotransferase; AST, aspartate transaminase; HDL-C, high-density lipoprotein cholesterol; LDL-C, low-density lipoprotein cholesterol; TC, total cholesterol; TG, triglyceride.
***P < 0.001 vs. CD group.
#P < 0.05, ##P < 0.01, ###P < 0.001 vs. FLD group.
Effects of maternal betaine supplementation on hepatic steatosis and inflammation in mice
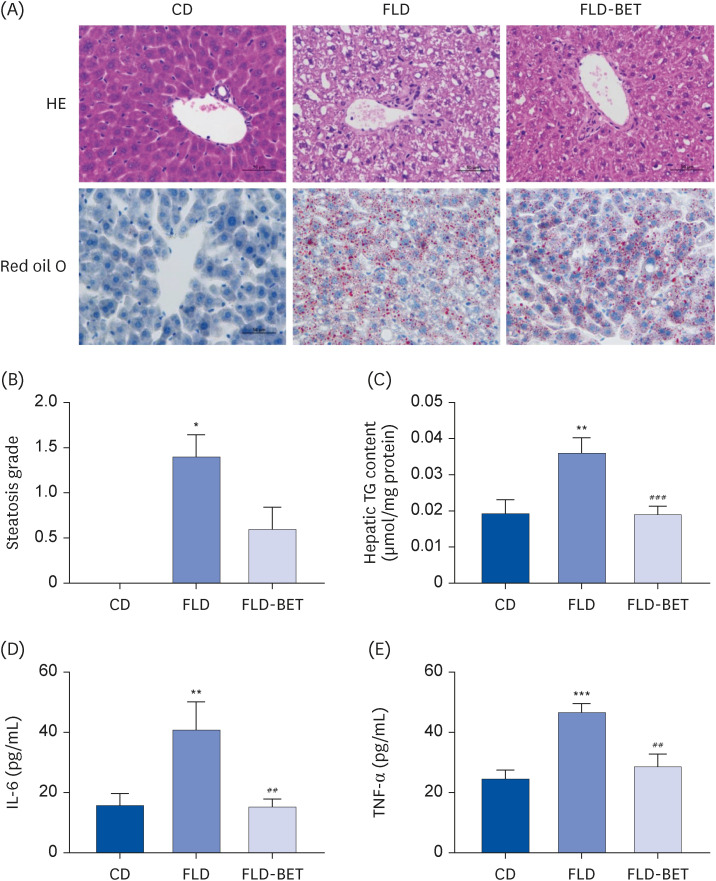
An analysis of HE-stained tissues from the offspring mice (Fig. 3A) showed that the FLD group exhibited obvious hepatic steatosis compared to the CD group. Oil red O staining (Fig. 3A) showed more severe lipid droplet formation in the FLD group than in the CD group. Moreover, the steatosis score was significantly higher in the FLD group than in the CD group (1.40 ± 0.24 vs. 0.00 ± 0.00, P = 0.012) (Fig. 3B). On the other hand, these changes were alleviated by maternal betaine supplementation. A significantly higher TG concentration was detected in the livers of the FLD group than in the CD group (0.04 ± 0.004 μm/mg vs. 0.02 ± 0.004 μm/mg, P = 0.005). Maternal betaine supplementation significantly reversed this TG accumulation (0.02 ± 0.002 μm/mg vs. 0.04 ± 0.004 μm/mg, P < 0.001) (Fig. 3C). The changes in histology and TG content were more pronounced in the livers of the dams (Supplementary Fig. 2). An analysis of the HE staining of liver sections showed significantly higher hepatic steatosis scores and inflammation scores in the dams of the FLD group (Supplementary Fig. 2). Although no significant difference in the hepatic inflammation score of offspring mice was found, the mice in the FLD group had significantly higher serum levels of IL-6 (40.83 ± 9.29 pg/ml vs. 15.84 ± 3.82 pg/mL, P = 0.005) and tumor necrosis factor-α (TNF-α) than those in the CD group (46.61 ± 2.90 pg/mL vs. 24.72 ± 2.80 pg/mL, P < 0.001). In contrast, maternal betaine supplementation reversed these changes (15.25 ± 2.58 pg/mL vs. 40.83 ± 9.29 pg/mL, P = 0.003 for IL-6; 28.64 ± 4.12 pg/mL vs. 46.61 ± 2.90 pg/mL, P = 0.001 for TNF-α) (Fig. 3D and E).
Fig. 3
Maternal betaine supplementation ameliorated hepatic steatosis in offspring mice exposed to high-fat diet and streptozotocin. (A) Liver section stained with HE and red oil O coloration. (B) Hepatic steatosis grade. (C) Hepatic TG concentration. (D) Serum IL-6 levels. (E) Serum TNF-α levels. Data are presented as the means ± SEs of the means.
HE, hematoxylin and eosin; CD, control diet; FLD, fatty liver disease; FLD-BET, fatty liver disease + 1% betaine; TG, triglyceride; IL, interleukin; TNF, tumor necrosis factor.
*P < 0.05, **P < 0.01, ***P < 0.001 vs. CD group.
##P < 0.01, ###P < 0.001 vs. FLD group.
Effects of maternal betaine supplementation on the expression of NLRP3 inflammasome in offspring mice
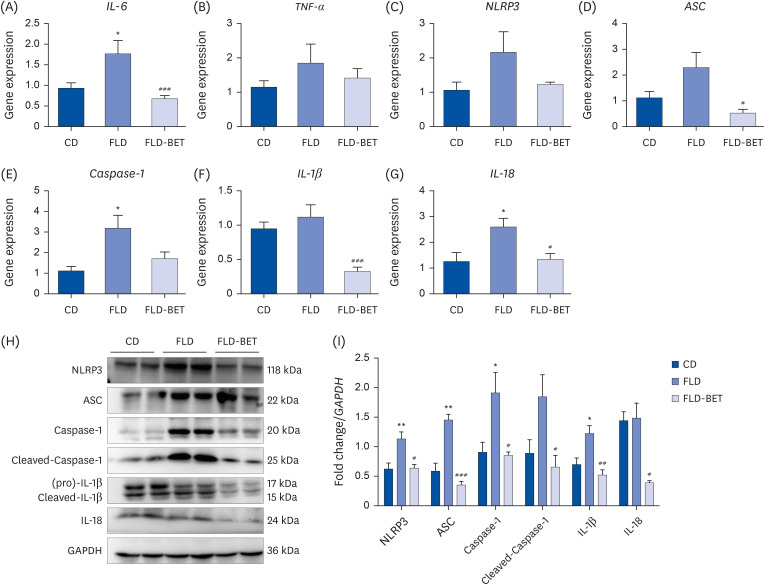
The mRNA expression of IL-6 (1.77 ± 0.32 vs. 0.94 ± 0.13, P = 0.012), Caspase-1 (3.20 ± 0.62 vs. 1.13 ± 0.20, P = 0.025), and IL-18 (2.59 ± 0.34 vs. 1.26 ± 0.34, P = 0.016) were significantly higher in the FLD group of offspring mice than in the CD group (Fig. 4A, E and G). After the intervention of maternal 1% betaine, the mRNA expression levels of IL-6 (0.68 ± 0.70 vs. 1.77 ± 0.32, P < 0.001), ASC (0.53 ± 0.14 vs. 2.30 ± 0.58, P = 0.049), IL-1β (0.33 ± 0.06 vs. 1.12 ± 0.18, P < 0.001), and IL-18 (1.34 ± 0.23 vs. 2.59 ± 0.34, P = 0.012) decreased significantly (Fig. 4A, D, F and G). Although the expression of NLRP3 mRNA did not reach statistical difference among the 3 groups (Fig. 4C), NLRP3 protein expression was significantly higher in the FLD group (1.14 ± 0.12 vs. 0.62 ± 0.10, P = 0.007) than in the CD group, which was reversed by maternal betaine supplementation (0.64 ± 0.06 vs. 1.14 ± 0.12, P = 0.011) (Fig. 4I). The protein expression of ASC (1.46 ± 0.09 vs. 0.59 ± 0.13, P = 0.001), Caspase-1 (1.92 ± 0.35 vs. 0.91 ± 0.17, P = 0.023), and IL-1β (1.23 ± 0.13 vs. 0.70 ± 0.11, P = 0.030) were significantly increased in the FLD group and decreased in the FLD-BET group (Fig. 4I). Maternal betaine intervention induced a decrease in the protein expression of IL-18 (0.39 ± 0.03 vs. 1.49 ± 0.25, P = 0.049) and Cleaved Caspase-1 (0.66 ± 0.20 vs. 1.85 ± 0.37, P = 0.039) (Fig. 4I).
Fig. 4
Maternal betaine supplementation inhibited the mRNA and protein expression of NLRP3 inflammasome in the liver of offspring mice exposed to high-fat diet and streptozotocin. (A-G) Hepatic mRNA expression of IL-6, TNF-α, NLRP3, ASC, Caspase-1, IL-1β and IL-18. (H) Representative image of western immunoblot analysis. (I) Hepatic protein expression of NLRP3, ASC, Caspase-1, IL-1β and IL-18. Data are presented as the means ± SEs of the means.
CD, control diet; FLD, fatty liver disease; FLD-BET, fatty liver disease + 1% betaine; IL, interleukin; TNF, tumor necrosis factor; NLRP3, nitric oxide dioxygenase-like receptor family pyrin domain containing 3; ASC, apoptosis-associated speck-like protein containing C-terminal caspase recruitment domain.
*P < 0.05, **P < 0.01 vs. CD group.
#P < 0.05, ##
P < 0.01, ###P < 0.001 vs. FLD group.
Effects of maternal betaine supplementation on hepatic DNA methylation level in offspring mice
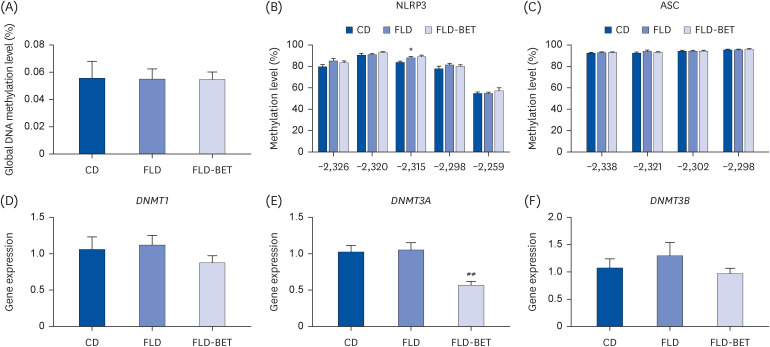
The global DNA methylation levels were similar in the liver of the 3 groups (Fig. 5A). The DNA methylation level at −2315 (88.29 ± 0.97 vs. 84.00 ± 0.93, P = 0.045) of NLRP3 in the FLD group was significantly higher than in the CD group. No significant difference in DNA methylation levels of NLRP3 was noted at other sites (Fig. 5B). The DNA methylation level of the ASC gene promoter region was similar at each site (Fig. 5C). The relative mRNA expression of DNA methyltransferase (DNMT)3A was lower in the FLD-BET groups than in the FLD group (0.56 ± 0.06 vs. 1.05 ± 0.09, P = 0.002). The mRNA expression DNMT1 and DNMT3B were similar in the 3 groups (Fig. 5D-F).
Fig. 5
Effects of maternal betaine supplementation on the hepatic DNA methylation levels in offspring mice. (A) Global DNA methylation level. (B, C) Methylation level of NLRP3 and ASC. (D-F) Gene expression of DNMT1, DNMT3A, and DNMT3B. Data are presented as the means ± SEs of the means.
CD, control diet; FLD, fatty liver disease; FLD-BET, fatty liver disease + 1% betaine; NLRP3, nitric oxide dioxygenase-like receptor family pyrin domain containing 3; ASC, apoptosis-associated speck-like protein containing C-terminal caspase recruitment domain.
*P < 0.05 vs. CD group.
##P < 0.01 vs. FLD group.
Effects of maternal betaine supplementation on hepatic one-carbon metabolism in offspring mice
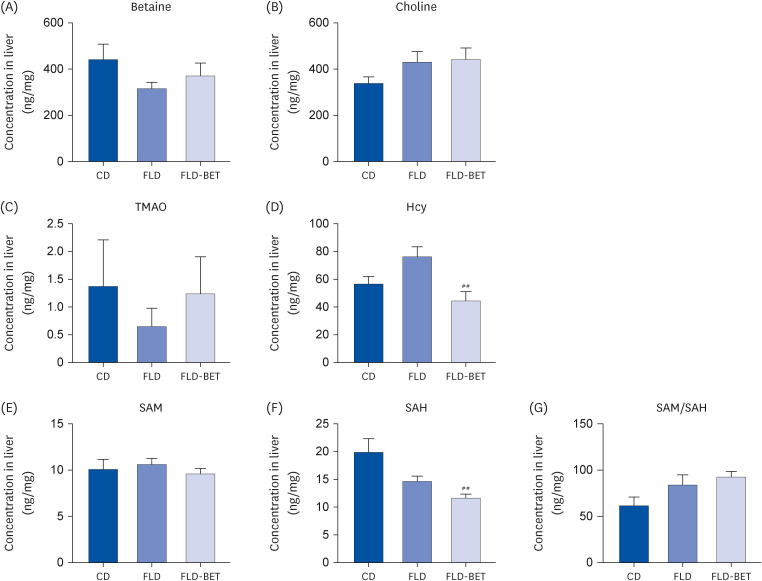
In the dams, HFD combined with STZ decreased the concentrations of betaine (1,651.34 ± 153.42 ng/mg vs. 3,142.37 ± 316.67 ng/mg, P = 0.012), choline (4,065.32 ± 408.26 ng/mg vs. 6,370.30 ± 335.10 ng/mg, P < 0.001), SAM (4.89 ± 0.21 ng/mg vs. 11.80 ± 1.26 ng/mg, P = 0.001), and SAM/SAH ratio (32.12 ± 2.75% vs. 66.92 ± 8.24%, P = 0.005) in the livers compared to CD group, while 1% betaine supplementation reversed the decreases in the betaine concentration (3,715.15 ± 350.02 ng/mg vs. 1,651.34 ± 153.42 ng/mg, P = 0.001), SAM concentration (6.59 ± 0.41 ng/mg vs. 4.84 ± 0.23 ng/mg, P = 0.004) and SAM/SAH ratio (54.69 ± 5.44% vs. 31.66 ± 2.21%, P = 0.018) induced by HFD and STZ (Supplementary Fig. 3). In offspring mice, maternal 1% betaine supplementation decreased the hepatic levels of Hcy (44.36 ± 6.35 ng/mg vs. 76.10 ± 6.84 ng/mg, P = 0.003) and SAH significantly (11.09 ± 0.67 vs. 14.70 ± 0.91 ng/mg, P = 0.008) (Fig. 6 D and F). In addition, the hepatic Hcy concentrations showed inverse relationships with the mRNA expression of TNF-α (r = 0.2616, P = 0.002), NLRP3 (r = 0.563, P = 0.002), ASC (r = 0.249, P = 0.041), and IL-18 (r = 0.268, P = 0.014). The hepatic IL-1β mRNA expression was inversely associated with the SAH concentration (r = 0.584, P = 0.002) and positively correlated with the SAM/SAH ratio (r = 0.442, P = 0.010).
Fig. 6
Regulation of maternal betaine supplementation on the hepatic levels of one-carbon metabolites in offspring mice. (A-G) Contents of betaine, choline, TMAO, Hcy, SAM, SAH, and SAM/SAH ratio in the livers of offspring. Data are presented as the means ± SEs of the means.
CD, control diet; FLD, fatty liver disease; FLD-BET, fatty liver disease + 1% betaine; TMAO, trimethylamine-n-oxide; Hcy, homocysteine; SAM, s-adenosine methionine; SAH, s-adenosine homocysteine.
##P < 0.01 vs. FLD group.
DISCUSSION
Previous studies have suggested that betaine supplementation in early life has a protective role in the fetal-original fatty liver induced by drug and HFD by rectifying the dysregulated genes involved in lipid metabolism [202122]. This study supports a new finding that betaine supplementation during pregnancy and lactation can downregulate the components of NLRP3 inflammasome and alleviate fatty liver in offspring mice induced by maternal HFD and STZ.
Several perinatal factors, including maternal nutritional status, obesity, and diabetes, can prime the offspring for later liver dysfunction and the development of NAFLD [423]. Hepatic TG accumulation already existed in the fetus (embryonic day 17.5) of dams fed a HFD for 6 weeks before gestation [21]. Consistent with these earlier findings, obvious lipid accumulation and steatosis in the livers of offspring mice were induced by maternal HFD and STZ. Maternal 1% betaine supplementation alleviated the NAFLD induced by HFD and STZ during pregnancy and lactation. With the development of steatosis, inflammatory cascades and pro-inflammatory cytokines were activated to contribute to the progression of SS to steatohepatitis [24]. Although the hepatic inflammation scores were similar in the groups of offspring mice, increased serum levels of proinflammatory cytokines, such as IL-6 and TNF-α, occurred before liver inflammation. The circulating levels of TNF-α and IL-6 are related to NAFLD, play a pathologic role, and can predict NASH development [25]. The increasing serum levels of TNF-α and IL-6 in offspring mice were corrected after maternal betaine intervention, which agrees with the effect of maternal betaine supplementation in glucocorticoid-induced NAFLD [22]. Therefore, increasing the betaine intake during pregnancy and lactation may be a potential intervention for preventing NAFLD.
Inflammasome is a major molecule that triggers pro-inflammatory cytokines maturation and production to initiate inflammation. Among these inflammasome families, NLRP3 inflammasome is the most extensively studied and well-characterized member. NLRP3 is a multiprotein complex that contributes to inflammation by activating Caspase-1, maturation, and the release of proinflammatory cytokines. In this study, maternal HFD and STZ intervention during pregnancy and lactation caused obvious NLRP3 and Caspase-1 activation, together with elevated the expression of ASC, IL-1β, and IL-18. Although the mRNA expression of NLRP3 showed an upward trend in the FLD group and a downward trend in the FLD-BET group, the differences were not significant. In addition, the protein of NLRP3 was significantly different among groups. This might be related to the transcription, processing, and degradation of mRNA, as well as the translation, localization, modification, and programmed disruption of proteins. Protein stably exists after translation, while translation appears to trigger mRNA decay [26]. In eukaryotes, the cellular concentrations of proteins correlate with the abundances of their corresponding mRNAs, but not strongly, and the correlation coefficient was only 0.40 [27]. The inhibition of the NLRP3 inflammasome is an effective method of improving NAFLD. Betaine has anti-inflammatory effects on several diseases by inhibiting NLRP3 inflammasome activation [13]. The betaine treatment inhibited the expression of NLRP3 components and reduced the levels of inflammatory cytokines, including IL-1β, IL-6, and IL-18, in the livers of fructose-induced NAFLD rats in a dose-dependent manner [14]. Betaine supplementation also inhibited the expression of the NLRP3 components and IL-1 in the livers of diabetes mice model [28]. In the present study, maternal betaine supplementation reversed the overexpression of NLRP3 and activation of the NLRP3 inflammasome in the livers of the offspring mice.
Hypomethylation in the promoter region is a feature of gene activation. A previous study on mycobacterium tuberculosis showed that NLRP3 activation was related to reducing the DNA methylation levels in the promoter region [29]. In the present study, no significant difference in the DNA methylation level was found in the promoter region of NLRP3-related genes among groups. On the other hand, the inhibiting effects of maternal betaine intake on NLRP3 inflammasome in the offspring mice may be related to its regulation of the one-carbon metabolism. Betaine is an intermediate one-carbon metabolism and can be derived endogenously from choline by several vital enzymes. Betaine provides a methyl group to convert Hcy into methionine, forming SAM. After providing the methyl group, SAM transforms into SAH. Maternal betaine supplementation during gestation and lactation had different regulations on the hepatic one-carbon metabolism between the dams and offspring. Maternal betaine supplementation increased the hepatic betaine, SAM contents, and SAM/SAH ratio in dams, which is in accordance with maternal betaine intake in HFD dams [21]. In the offspring mice, reduced Hcy and SAH concentrations were detected in the livers of the betaine-exposed offspring mice. Several disease models have been used to examine the role of NLRP3 inflammasome activation in Hcy-induced inflammation [3031]. Hcy contains a reactive sulfhydryl group and can be auto-oxidized with another Hcy molecule to generate various disulfides (Hcy–Hcy, Hcy–Cysteine, Hcy–Glutathione), resulting in the release of reactive oxygen species [32]. Oxidative stress triggers the assembly of NLRP3, ASC1, and pro-Caspase-1 to form a multiprotein complex [33]. A significantly reverse association was observed between the Hcy concentration and expression of NLRP3 inflammasomes. Therefore, the inhibited activation of NLRP3 inflammasome by maternal betaine supplementation in the livers of offspring mice may be related to its regulation on Hcy.
The prevalence of NAFLD is increasing with the growing global obesity epidemic, and the people affected are becoming younger. Therefore, it is essential to explore ways of safe nutritional interventions to prevent NAFLD early in life. The present study showed that maternal betaine supplementation during pregnancy and lactation alleviates fetal-derived NAFLD induced by HFD combined with STZ. The underlying mechanism may be related to inhibiting the activation of NLRP3 inflammasome.
 XML Download
XML Download