

 PDF
PDF Citation
Citation Print
Print


INTRODUCTION
Cisplatin is a widely used chemotherapeutic agent for treating cancer patients [1-3]. Its bioactivity is based on disruption of calcium homeostasis, lipid peroxidation, and DNA damage to induce apoptosis of cancer cells [4]. Many researchers and studies have generally been accepted that cisplatin is one of the most successful anti-cancer agents, and its therapeutic benefits have been well documented. Nevertheless, there is limiting considerably its prolonged utilization because leads to a range of acute and late toxicities [5,6].
Cisplatin-induced toxicities lead to numerous side effects including cardiovascular and metabolic diseases, hypogonadism, poor mental health-related quality of life, peripheral neuropathy, and cognitive impairment [7]. Particular, cognitive impairment is recognized as the most typical symptom induced by cisplatin neurotoxicity in the central nervous system (CNS). Cisplatin-induced cognitive impairment is known as chemo-brain, and also frequently reported by patients treated for tumor outside the CNS [8]. Actually, it has been reported that approximately 60%–75% of patients suffer from both short- and long-term cognitive impairment during and following cisplatin treatment. Along with, some patients have psychological issues such as depression, anxiety, and lethargy [9]. Importantly, the current problem is to note that these health concerns may lead to future treatment refusal and/or reduce the effectiveness of advanced-stage treatment.
Although the pathogenesis of cisplatin-induced cognitive impairment is not completely understood, accumulating evidence suggests that it is similar to biological progressive brain aging and that cisplatin contributes to both structural and functional changes in the hippocampus causing neuropathologic alterations [10]. The hippocampus plays a pivotal role in cognitive function including attention, memory, processing speed, and learning. However, the hippocampus is very vulnerable and sensitive to anti-cancer drugs such as cisplatin [11], and dysfunction of it strongly is implicated in to cause and/or consequence of various pathological conditions. It has been reported that hippocampal dysfunction is associated with an increased prevalence of mild cognitive impairment and neurodegenerative disorders such as Alzheimer’s disease and dementia [12].
Systemic inflammation is recognized as a critical predictor the onset of the various disease in clinical and animal studies. Among with many pathways, increased inflammation plays a vital role in the etiology of neurodegenerative diseases, and it has been well documented [13,14]. Many studies have reported that cisplatin accelerates releases of pro-inflammatory cytokines such as tumor necrosis factor alpha (TNF-α) and interleukins (ILs), which are involved in oxidative stress, mitochondrial damage, neuronal apoptosis, dysregulation of neurotransmitters, and inhibition of neurogenesis in the hippocampus [15,16]. In addition, synaptic dysfunction such as synapses loss and deficits were thought to be strongly associated with cognitive impairment. Some studies reported that increased inflammatory factors in the CNS contributes to weaken target neuron activity through modification of the translational expression of proteins that control synaptic stability and neuronal excitability [17,18]. In this regard, it has been demonstrated that increased inflammatory response by cisplatin is involved in the regulation of synapsin-1 and -2 which are essential for synaptic plasticity and cognition [19].
Given that increased inflammation is a hallmark of cancer pathophysiology [20] and that inflammation response after cisplatin treatment, modulation of specific cytokines may be a potential strategy to prevent and mitigate cognitive impairment caused by cisplatin. Recently, there has been a growing body of research on pathophysiological mechanisms of cisplatin-induced cognitive impairment, but effective early intervention strategies to delay or ameliorate has not been developed yet.
Exercise is widely accepted as an effective non-pharmacological intervention to prevent and improve various diseases and to maintain health. Many clinical and animal studies indicate that exercise contribute to alteration to brain activation and/or damage. Recent studies reports that well-organized regular exercise and physical activity bring benefits to cisplatin toxicities-mediated side effects in the peripheral regions including skeletal muscle, renal, and heart [21-23] and emphasize the importance of consistent of exercise practice to potential protective effects. In contrast to, there is a limited amount of research on the effects of exercise on common CNS outcomes caused by chemotherapy. Given the potential for cisplatin-induced cognitive impairment, this study aimed to determine the efficacy of exercise in protecting against early indicators of cognitive impairment mediated by hippocampal damage following cisplatin treatment.
METHODS
Experiment animals
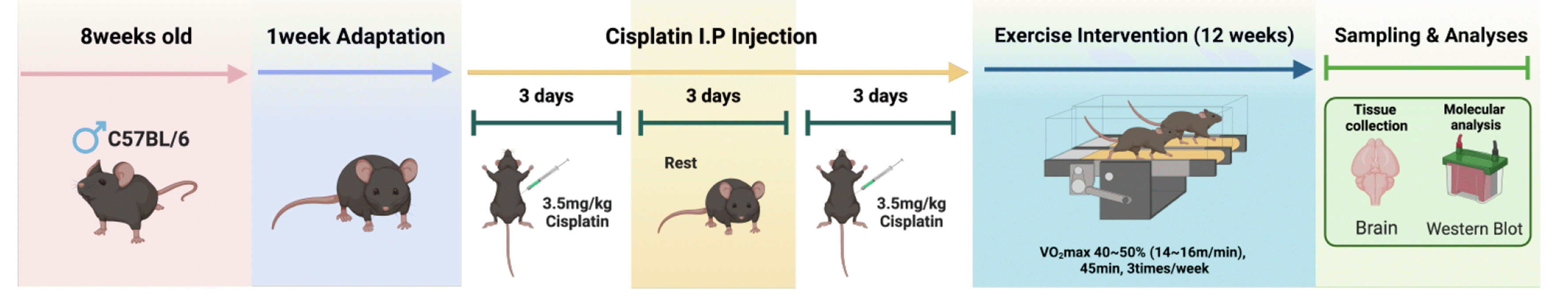
Eight-week-old C57BL6 mice were purchased from the Yeungnam Bio Company. The mice were randomly divided into three groups as follows: control (CON, n = 8); cisplatin injection (Cis-CON, n = 8); exercise with cisplatin injection (Cis-EXE, n = 8). The animals were preserved at a temperature of 20°C ± 2.5°C under a 12-h light-dark cycle with free access to food and water. All experimental procedures were approved by the Institutional Animal USE and Care Committee of Inje University (2023-006).
Cisplatin injection and experimental protocols
To induce hippocampus damage by cisplatin administration, we used the platinum-based compound cisplatin (Sigma-Aldrich, Cat# 232120), which has been reported to persist long-term for about 2–5 weeks. Male mouse at the age of 4–5 months were treated with daily cisplatin (3.5 mg/kg/day) through intraperitoneal injection. Notably, previous studies have reported that cisplatin penetrates the blood-brain barrier and persists long-term for about 2–5 weeks after its administration and this dose causes significant neurotoxicity to hippocampal neurons, leading to cognitive impairment in rodents [24]. The overall experimental procedure is shown in Fig. 1.
Aerobic exercise protocol
The Cis-EXE group was familiarized with treadmill running (1050-RM, Exer-3/6 treadmill; Columbus Instruments) three times per week at a pace of 10 m/min for 15 min. During the first week after familiarization, the mice exercised by treadmill running at 12 m/min on a 0% gradient for 30 min/day. The exercise intensity was gradually increased to 14–16 m/min (VO2max 40%–50%) on a 0% gradient for 30–45 min/day over 12 weeks [25].
Tissue preparation and Western blotting
Tissue samples were collected from the hippocampus, frozen on ice, and stored at –80°C until analysis. Samples were homogenized using RIPA buffer. Samples were spun at 14,000 rpm for 15 min at 4°C, and total protein concentration of the supernatant was determined by a Bradford assay. Tissues were collected and dissolved in a radioimmunoprecipitation analysis buffer using a cocktail of protease inhibitors. Protein lysate was electrophoresed with sodium dodecyl sulfate-polyacrylamide gel and transferred to nitrocellulose membranes. Next, 5% bovine serum albumin was used to block the membranes at 4°C for 2 h. The membranes were subsequently incubated with the following primary antibodies at 4°C overnight: synapsin-1 (1:1,000, Cell Signaling), synapsin-2 (1:1,000, Cell Signaling), TNF-α (1:2,000, Cell Signaling), IL-6 (1:2,000, Cell Signaling), IL-1β (1:2,000, Cell Signaling), IL-10 (1:2,000, Cell Signaling), super oxide dismutase (SOD) (1:1,000, Abcam), Bcl-2 (1:1,000, Abcam), Bax (1:1,000, Abcam) and the reference protein GAPDH (1:100,000, Cell Signaling). After washing with Tris-buffered saline with Tween 20, the membranes were incubated with HRP-conjugated secondary antibody (anti-mouse or anti-rabbit, Cell Signaling) at a dilution of 1:2,000 at 4°C for 4 h. An enhanced chemiluminescence kit (Merck Millipore) and imaging system (Bio Spectrum 600; UVP) were used to visualize the bands. ImageJ (National Institutes of Health) was used to quantify the blots.
Statistics
Statistical analyses were performed using GraphPad Prism version 8.1.0. software (GraphPad, Inc.). All data are presented as the mean ± standard deviation. Different groups were evaluated using one-way analysis of variance and Tukey’s honestly significant difference test for post-hoc analysis. A p-value of ≤ 0.05 was considered to indicate a statistically significant difference.
RESULTS
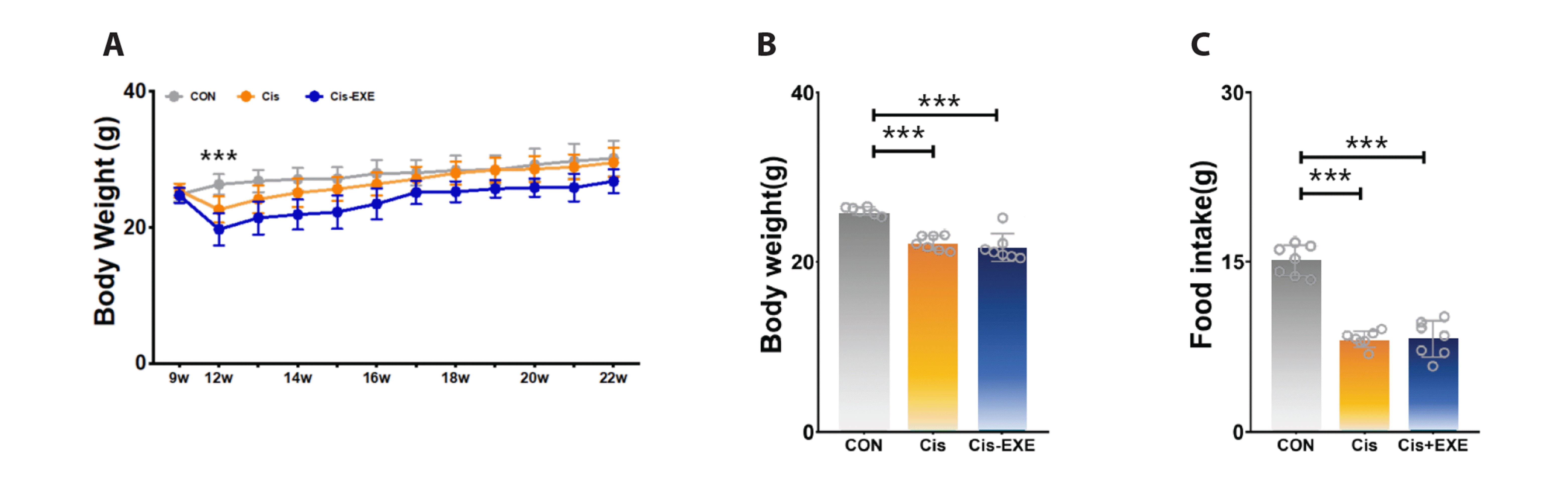
Cisplatin toxicity induces decrease of body weight and food intake
To validate the effects of cisplatin toxicity, we measured body weight and food intake among the treatment groups (Fig. 2). Compared to the CON group, body weight and average food intake (g/week) were significantly decreased in the Cis-CON and Cis-EXE groups. Our data pointed to the fact that body weight loss and decreased appetite are the most commonly reported initial adverse effects following chemotherapy and these results are consistent with previous studies indicative of successful induction of cisplatin toxicity.
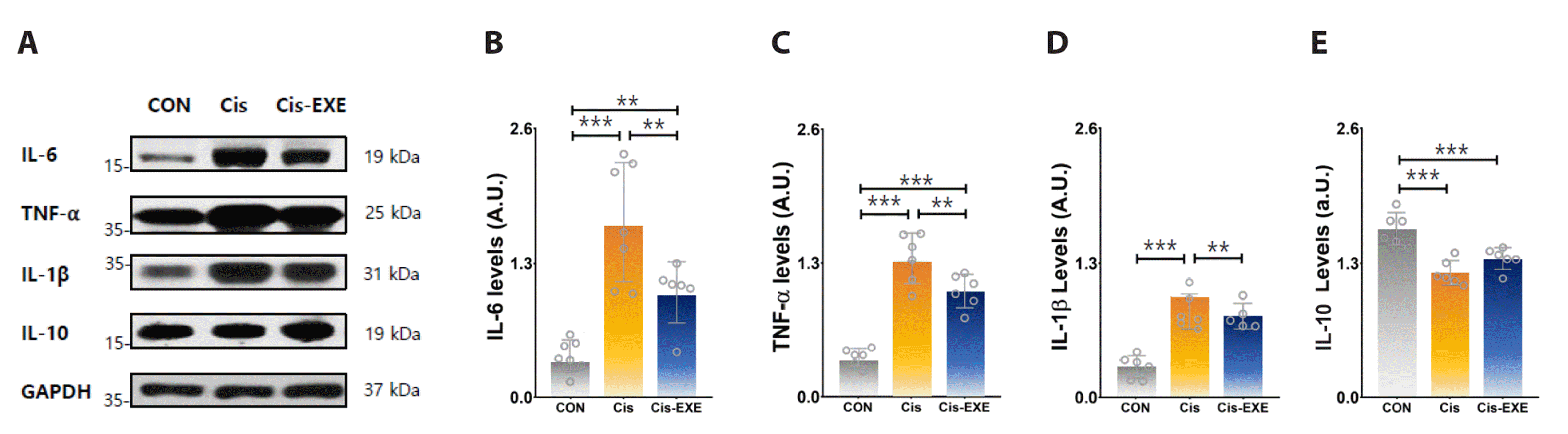
Exercise has protective effects on the cisplatin-induced neuroinflammation in the hippocampus
Since the inflammatory response is involved in the pathological process of cognitive impairment, we used western-blot to observe the alteration of IL-6, IL-1β, TNF-α, and IL-10. The IL-6, IL-1β, and TNF-α levels were elevated in the Cis-CON group when compared with the CON group, as shown in Fig. 3. Compared to the Cis-CON group, there was a significantly decreased levels of IL-6 and TNF-α in the Cis-EXE group. Our data indicated that immediate exercise intervention after cisplatin treatment may contribute to the inhibition of their expression. The level of IL-10 was slightly increased in the Cis-EXE group compared to the Cis-CON group, but the difference between the two group was not significant.
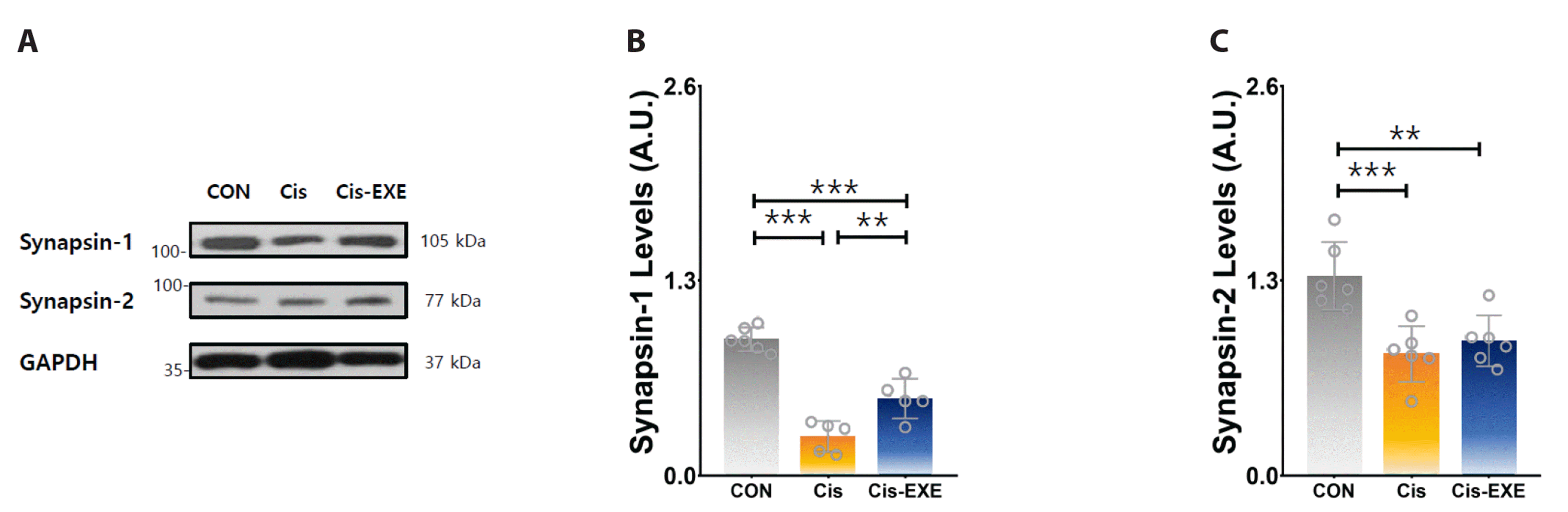
Exercise improves the expression of synapse-related proteins
To clarify the differences in the expression of synapse-related protein level among the groups, the protein expression levels of synapsin-1 and -2 in hippocampus tissue were determined (Fig. 4). The protein expression levels of synapsin-1 and synapsin-2 were significantly lower in the Cis-CON group compared with the CON group. Exercise increased the expression levels of synapsin-1 effectively. Synapsin-2 was no significant difference between Cis-CON and Cis-EXE group. These findings suggested that exercise alleviates some synapse-related factors in the hippocampus of mice with cisplatin injection.
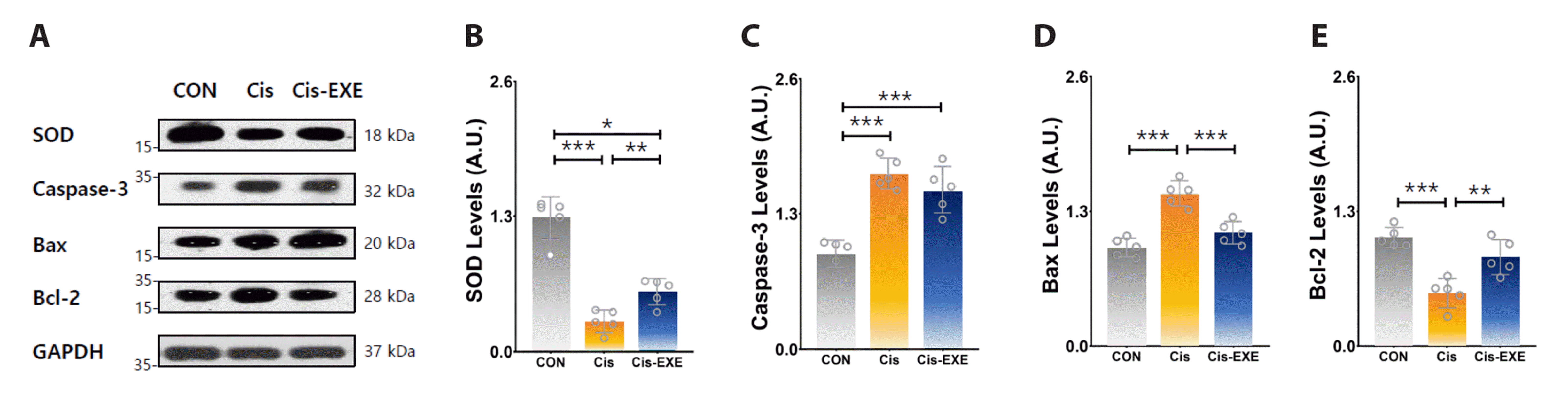
Exercise improves expression of antioxidant enzyme and pro-apoptosis protein
To clarify the differences in the expression of antioxidant enzyme and apoptosis-related protein level among the groups, the protein expression levels of SOD, Bax, Bcl-2, and caspase-3 in hippocampus tissue were determined (Fig. 5). Compared to the Cis-CON group, the SOD and Bcl-2 protein levels were significantly increased and the Bax and caspase-3 protein levels were significantly decreased in the Cis-EXE group. These findings indicated that exercise may have antioxidant and anti-apoptosis effects in cisplatin-mediated hippocampal damage.
DISCUSSION
To the best of our knowledge, this study is the first to elucidate the beneficial effects of exercise as a non-pharmacological strategy to control cisplatin-induced cognitive impairment. Our results suggest that regular exercise intervention could be a viable option to mitigate cisplatin-induced neurotoxicity, which might be attributed to the significant decrease in some inflammatory and increase synapse- related factors.
Recently, many studies have focused on effective pharmaceutical strategies to control the chemotherapy-accompanied various side effects. However, previous results have been far from satisfactory and there may be a need for more effective treatment options or complementary therapies. Over the past decades, it has been demonstrated that exercise plays an important role in alleviating cognitive impairment mediated by certain factors such as aging and metabolic disorders. This indicates that exercise interventions have the potential to improve the cognitive deficits associated with chemotherapy. Regular exercise intervention could be a safe strategy for managing cognitive impairment and mental health problems.
To validate the successful induction of cisplatin toxicity, we observed body weight and food intake. Previous studies have reported that cytotoxic agents, including cisplatin, can cause weight loss despite their anticancer effects, and that cisplatin-mediated toxicity contributes to lipid catabolism and muscle wasting [26,27]. Compared to the CON group, body weight and average food intake (g/week) were significantly decreased in the Cis-CON and Cis-EXE groups. However, the final body weight was no significant difference between the Cis-CON and Cis-EXE groups. These results are consistent with previous studies showing that the most common symptoms of cisplatin toxicity are loss of body weight and appetite [28].
Chemotherapy-induced cognitive impairment results from widespread damage to the CNS caused by neurotoxicity [29]. Increased inflammation is the primary characteristic of cisplatin-mediated neurotoxicity, which contributes to short- or long-term cognitive impairment. Cisplatin administration can cross the blood-brain barrier and directly damage the hippocampus. Accumulating evidence suggests that inflammatory pathways may play a critical role in the underlying mechanisms of cognitive decline and cisplatin amplifies the release of inflammatory factors that trigger a cascade of cellular damage and alter neurochemistry by enhancing the release neurotransmitters in hippocampus [30,31]. In this regard, it has been demonstrated that TNF-α, IL-6, and IL-1β are prominent pro-inflammatory factor observed in the hippocampus of cognitive impairment models, and activation of these may directly contribute to cognitive impairment [32]. Inversely, IL-10 acts as anti-inflammatory and thus suppresses the activity of pro-inflammatory mediators. In the current study, we observed a significant increase in IL-6, IL-1β and TNF-α levels in the Cis-CON group compared to the CON group. Conversely, the exercise group showed a significant decrease in the aforementioned cytokines, while IL-10 showed a significant increase. Persistent these cytokines expression activates astrocytes and microglia for a strong inflammatory response and subsequently may then serve as predictors for the development of cognitive impairment [33,34]. De Senna et al. [35] and Chupel et al. [36] demonstrated that exercise improves the level of inflammation in brain by safeguarding the structural integrity of the blood-brain barrier, which eventually leads to reduction of TNF-α and IL-6 expression. Our results are in line with clinical and animal studies [37,38] that have identified elevated levels of pro-inflammatory factors as a significant predictor of a wide range of neurodegeneration disorders and observed that exercise has anti-inflammatory effects.
In addition, synaptic plasticity is the major cellular mechanism underlying cognitive function including learning and memory and is considered a key function in the process of systems memory consolidation. An increase in cytokines and free radicals causes synaptic dysfunction and synaptic integrity, which are recognized as initial signs of neurodegeneration including cognitive decline [39]. Synapsins are known as representative synaptic plasticity-related proteins and are abundant in most nerve cells. Synapsin-1 and -2 are modulated neurotransmitter release at the pre-synaptic terminal and acts as a link between extracellular stimuli and the intracellular signaling events activated upon neuronal stimulation [40]. Previous studies demonstrated that patterns of synapsins protein expression are closely associated with neurological phenotypes. A significant decrease of the synapsin-1 level in the hippocampus is predominant in the early phase of the development of cognitive deficits and Alzheimer’s disease [41,42]. Moreover, overall synapsin deletion can lead to behavioral alterations similar to neuropsychiatric conditions [43]. These results indicate that interventions focused on maintaining the expression of synapsins in the hippocampus may be beneficial for individuals at a delaying the onset of cognitive impairment. In the current study, we observed that only synapsin-1 was significantly increased in the Cis-EXE group compared with the Cis-CON group. In this regard, it has been reported that synapse-related proteins depended on inflammatory response and the increase of cytokines such as TNF-α and IL-6 inhibits the expression of synapsin-1 and -2 in normal biological aging [44]. On the other hand, many studies indicated that long-term exercise might protect hippocampal by enhancing synaptic connections and either promotes synaptic formation and/or maintain synapses that would have otherwise alteration after brain damage [38,45]. Eventually, reduced inflammation and increased plasticity within hippocampus are thought to be crucial for cognitive function [46,47]. According to previous findings, it have been demonstrated that exercise significantly decreased microgliosis, TNF-α, and IL-1β in the CA1 area of hippocampus and that long-term exercise contributes to protect hippocampal synapses and exercise-mediated FNDC5/irisin improves synaptic plasticity in mice [48,49].
Along with this, our results demonstrated the role of exercise in antioxidant and anti-apoptosis effects. Our data shows that exercise-induced antioxidant activity offsets apoptosis in the hippocampus damage mediated by cisplatin. Exercise is well known as an effective method to maintain and improve the physiological function of overall brain regions. In previous studies, Zare et al. [50] reported neuroprotective effects of exercise mediated by inhibition of apoptosis in the hippocampus and Navazani et al. [51] suggested that exercise decreased the expression of BAX and increased the expression of Bcl-2 in the hippocampus of rats treated with thimerosal, which causes neurotoxicity. Exercise plays a crucial role for controlling activation of antioxidant enzyme system. It was reported that regular exercise-mediated activation of antioxidant enzyme has protective effects on inflammatory and oxidative stress in the hippocampus [52,53].
Although the pathogenesis of cisplatin-induced cognitive impairment is not fully understood, previous studies have shown that the disruption of inflammatory signaling pathways and synaptic plasticity are involved in onset of mild cognitive impairment and neurodegenerative disorders by causing neurotoxicity. Given that, it is believed that significant control in inflammation and synaptic-related proteins through regular exercise intervention from the early phase chemotherapy may contribute to preventing and improving the risk of brain damage-accompanied cognitive impairment. Our study has certain limitations. We have not conducted behavioral tests to assess cognitive dysfunction. The previous studies using cognitive behavioral assessments have documented increased inflammation as a significant factor in cognitive deficits. This study only focused on the effects of exercise as a pre-intervention strategy on the toxicity of cisplatin, regardless of tumor control because the toxicities of it affect not only target tumor cells but also normal host cell.
It is essential to understand the hippocampus damage mediated by cisplatin and to develop therapeutic intervention strategies in order to improve the quality of life of cancer patients. Exercise plays an important role in cognitive functions, regardless of the presence or absence of disease. Our study suggests that engaging in exercise immediately after receiving cisplatin treatment could be a meaningful approach to prevent or improve cisplatin-induced cognitive impairment by regulating inflammation and synapse-related factors.
 XML Download
XML Download