

 PDF
PDF Citation
Citation Print
Print


INTRODUCTION
Cholesterol is a crucial component of cellular membranes, constituting approximately 20% of all membrane lipids. It plays a vital role in maintaining integrity and fluidity of cell membrane and cellular physiology [1]. However, cholesterol is susceptible to oxidation, resulting in the formation of oxysterols. Among oxysterols, 27-hydroxycholesterol (27OHChol), which is generated either by autooxidation or mitochondrial cytochrome P450 enzyme sterol 27-hydroxylase (CYP27A1), stands out as the most abundant oxysterol in the bloodstream and peripheral tissues [2-4]. It is a bioactive metabolite with remarkable implications for various biological processes, including cholesterol homeostasis, lipid metabolism, gene expression, and innate immune response [5-7].
27OHChol exerts pleiotropic effects on monocytic cells, inducing the differentiation and polarization of monocytes and macrophages into mature dendritic cell phenotype and pro-inflammatory M1 type, respectively [8,9]. Activated M1 monocytic cells display increased expression of cytokines, chemokines, and surface molecules related to inflammatory responses [7,10,11]. Given the association of M1 monocytic cell accumulation and oxysterol dysregulation with conditions such as atherosclerosis, adiposity and obesity [12-15], ongoing research seeks to explore the intricate mechanisms and implications of 27OHChol in human inflammatory diseases.
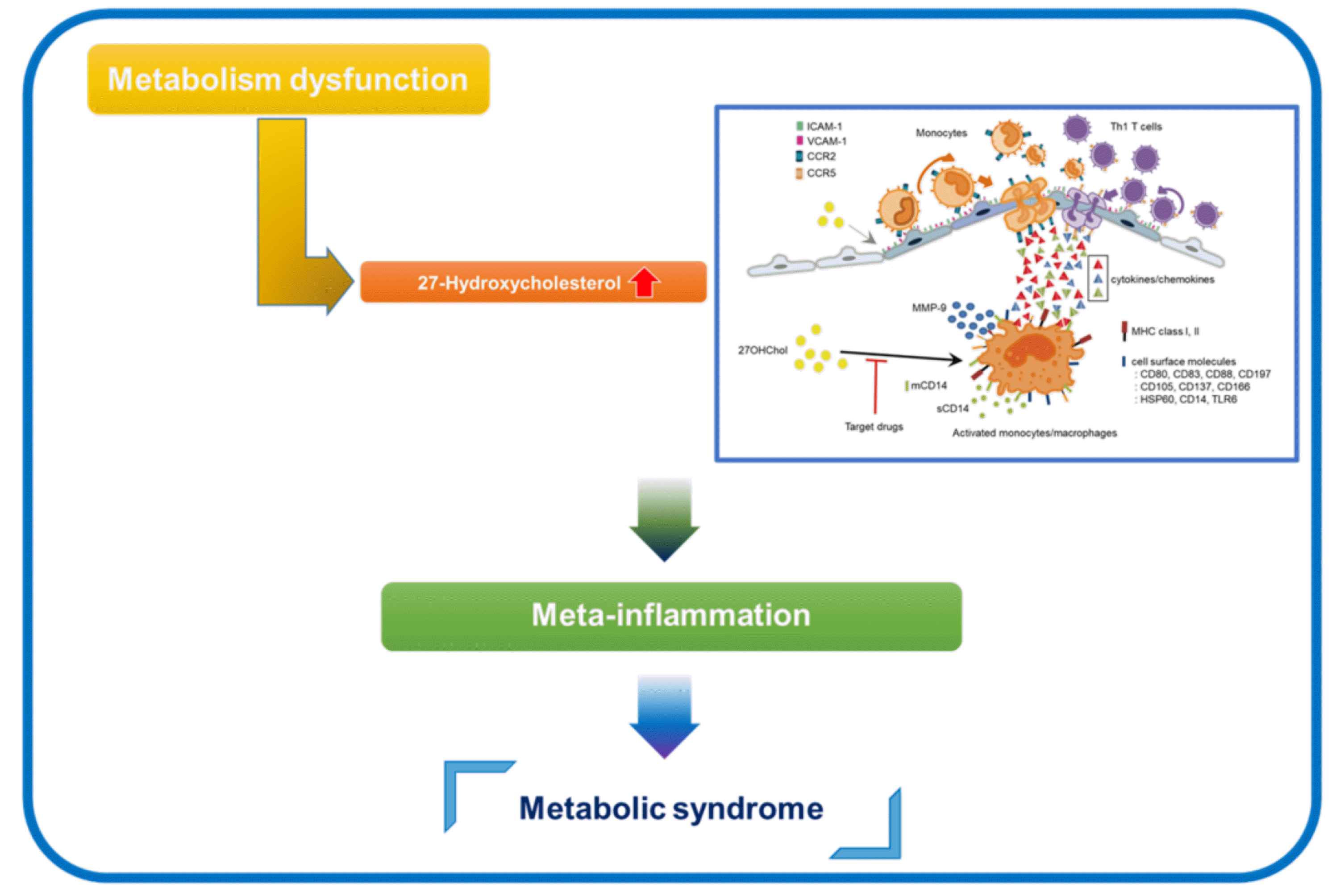
Metabolic inflammation, also known as meta-inflammation, is characterized by low-grade, chronic inflammation in metabolic disorders like cardiovascular diseases, obesity, and diabetes [16]. It differs substantially from acute inflammation regarding underlying causes, duration, and immune cell activation. While acute inflammation typically results from external factors like toxin exposure, microbial invasion or physical injury, meta-inflammation primarily stems from metabolic abnormalities [17]. Acute inflammation is transient, lasting for a short period, usually hours to days, while meta-inflammation persists for months or even years [18]. Meta-inflammation involves complex interactions among metabolic abnormalities, immune cells, and organs. It is featured by activation of monocytes/macrophages and release of pro-inflammatory cytokines in response to metabolic stress. These events lead to immune cell infiltration into affected tissues and dysregulated signaling pathways associated with development and progression of metabolic syndrome [16,18,19]. Therefore, understanding the unique characteristics of metabolic inflammation is pivotal for developing targeted approaches to manage and treat chronic metabolic disorders.
The objective of this review article is to highlight the involvement of 27OHChol in the pathophysiology of human diseases, particularly in the context of meta-inflammation. We presume that the activation of monocytic cells by 27OHChol can trigger and amplify inflammation, contributing to the progression and complications of major chronic diseases. By investigating the role of 27OHChol in meta-inflammation, we aim to gain insights into its part in the pathogenesis and treatment of chronic human diseases.
27OHCHOL AND CYTOKINES/CHEMOKINES
Cytokine and chemokine contribute to the establishment of chronic low-grade inflammation alongside with other inflammatory mediators [16,19]. 27OHChol enhances expression of both anti- and pro-inflammatory mediators in monocytic cells. However, it was noteworthy that the pro-inflammatory M1 molecules exhibit a significantly stronger expression, leading to an overall inflammatory response [8]. Studies from our laboratory have reported that cytokine and chemokines like CCL2, CCL3, CCL4, CXCL8, and tumor necrosis factor (TNF)-α are involved in 27OHChol-induced inflammatory responses.
27OHChol induces CCL2 secretion by liver X receptor-independent and Akt-dependent mechanisms, which indicates involvement of multiple pathways in CCL2 expression [20,21]. CCL2 binds to its receptor, C-C motif chemokine receptor-2 (CCR2), and recruits immune cells, particularly monocytes and macrophages, to sites of inflammation [22,23]. In agreement with the fact, CCL2 secreted from the 27OHChol-treated cells enhances monocytic cell migration through a CCR2-dependent mechanism [10,21]. Once recruited at the site of inflammation, monocytes differentiate into macrophages which are activated by pro-inflammatory molecules. These activated macrophages, in turn, release chemokines that attract more monocytes [24]. Unless the stimulus is removed, this vicious cycle of inflammatory response can persist for years, contributing to chronic low-grade inflammation. The monocyte- and macrophage-mediated response is central to the pathogenesis of obesity and atherosclerosis [18,24,25]. Therefore, the accumulation of 27OHChol in vasculature and adipose tissue can result in an increased number of monocytes and macrophages within the tissues via the CCL2/CCR2 axis, thereby promoting vascular and adipose tissue inflammation [12,15].
TNF-α is one of cytokines whose production is enhanced following exposure of monocytic cells to 27OHChol [26]. This cytokine influences adipose tissue inflammation, glucose metabolism, and cardiovascular remodeling through tissue-specific mechanisms [27]. TNF-α stimulates its own production within adipose tissue, creating a positive feedback loop that sustains inflammation, which perpetuates the pro-inflammatory environment in obesity [28]. TNF-α also directly impacts adipocyte function and metabolism since it impairs adipogenesis, resulting in adipocyte hypertrophy (enlarged fat cells) and adipose tissue dysfunction [28]. In addition, it interferes with adipocyte insulin signaling [29]. In the vasculature, TNF-α alters endothelial function, affecting the interaction between endothelial cells and blood cells and leading to vascular dysfunction [30], and increases the transcytosis of low-density lipoprotein (LDL) across endothelial cells, facilitating the accumulation of LDL in the subendothelial space of vessel walls and promoting early atherosclerosis [31]. Moreover, released TNF-α from tissues into the bloodstream can have systemic effects [27,32].
27OHCHol induces the expression of CXCL8/interleukin (IL)-8 in monocytic cells via CD88 [33]. CXCL8 plays a vital role in recruiting and activating neutrophils, immune cells involved in the early stages of host defense against infection and injury [34]. CXCL8 is implicated in the pathogenesis of atherosclerosis [35]. In the context of metabolic inflammation, upregulated CXCL8 expression has been observed in adipose tissue [36]. The increased expression of CXCL8 under conditions rich in 27OHChol may cause the recruitment and activation of immune cells that promote tissue damage.
27OHChol increases the production of CCL3 and CCL4 that preferentially enhance migration of T cells expressing the C-C chemokine receptor 5 (CCR5) [11]. CCR5, characteristic of Th1 T lymphocytes, is expressed at high levels on the surface of Th1 T cells [37]. In the process of 27OHChol-induced inflammatory response, Th1 cell movement increases towards the sites of inflammation, leading to Th1 dominant conditions. Th1 T cells induce further inflammation by secreting cytokines including interferon-gamma (IFN-γ) and TNF-α [38]. IFN-γ enhances activation of macrophages, aids foam cell formation, and causes destabilization of atherosclerotic plaques in the vasculature [39-41]. In adipose tissue, cytokines secreted from Th1 cells sustain local inflammatory responses by activating other immune cells, and therefore enhance adipose tissue inflammation and contribute to the development of metabolic diseases [42,43].
The role of Th1 T cells in type 2 diabetes is an area of active research. An imbalanced Th1/Th2 cytokine profile has been implicated in disease development and progression. Elevated Th1/Th2 cytokine ratios, such as IFN-γ/IL-5 and IL-2/IL-5, are correlated with type 2 diabetes and its complications, including retinopathy and cardiovascular issues [44]. This result suggests that an exaggerated Th1 response coupled with a relative decrease in the Th2 response may contribute to the pathogenesis of type 2 diabetes in a milieu rich in 27OHChol.
27OHCHOL AND PATTERN RECOGNITION RECEPTORS
27OHChol affects the expression of molecules on the cell surface in addition to secretion of cytokines and chemokines. Previous studies from our laboratory demonstrated that 27OHChol upregulates the levels of heat shock protein 60 and multiple CD molecules including CD80, CD83, CD88, CD105, CD137, and CD166 [9,45,46]. These increases in expression are important indicators because the individual molecules are involved in cell characteristics, differentiation, and function. 27OHChol also elevates levels of pattern recognition receptors (PRRs), like Toll-like receptor 6 (TLR6) and CD14 [10,47]. Considering the results of previous studies from our laboratory, 27OHChol seems to prime monocytic cells by upregulating the PRRs such that their metabolic ligands can trigger a cascade of signaling events enhancing inflammation.
27OHCHol upregulates TLR6 expression on cell surface, and the 27OHChol-activated monocytic cells secrete IL-1α in response to a TLR6 ligand [47]. Upon release, IL-1α binds to its receptor, leading to an innate immune response that activates PRRs due to IL-1α's similarity to infectious pathogens. Therefore, IL-1α promotes inflammation by inducing the production of other cytokines, chemokines, and small-molecule mediators [48]. This results in the systemic elevation of pro-inflammatory cytokines and transient reactions [49].
Monocytic cells activated with 27OHChol exhibit both increased expression of membrane-bound CD14 and enhanced secretion of soluble CD14 [20,45]. CD14 is crucial for the recognition of and response to lipopolysaccharide (LPS) [50]. The stimulation of monocytic cells with LPS in the presence of 27OHChol results in a super-induction of CCL2 compared with LPS alone [10], indicating that 27OHChol further enhances the CD14-mediated inflammatory pathway. However, the CD14 expression coupled with exaggerated LPS response and 27OHChol-induced inflammation were suppressed by treatment with anti-inflammatory drugs as well as HSP90 inhibitors [8,51-53]. CD14 also recognizes and binds various ligands associated with metabolic disorders. These ligands comprise oxidized LDL, a key contributor to atherosclerosis, and saturated fatty acids like palmitate and stearate that are elevated in obesity [54,55]. The CD14-mediated recognition of saturated fatty acids and other metabolic ligands promotes chronic low-grade inflammation and thereby can contribute to the pathogenesis and progression of insulin resistance, type 2 diabetes, and obesity [55-57]. Taken together, these findings suggest that 27OHChol is thought to be a key cholesterol metabolite involving CD14 in meta-inflammation.
CONCLUSION AND PROSPECTS
It has become evident that 27OHChol plays a key role in meta-inflammation, rendering it a pivotal factor in the development and progression of chronic diseases. This role is primarily attributed to its capacity to activate monocytes and macrophages and induce their expression of pro-inflammatory cytokines and chemokines, thereby disrupting metabolic signaling pathways (Fig. 1). Therefore, gaining a comprehensive understanding of the mechanisms and functions of 27OHChol in meta-inflammation is essential for the development of targeted approaches to manage and treat chronic metabolic disorders. We consider that the targeting of pathways involved in 27OHChol-induced inflammation may hold therapeutic promise for the new treatment of chronic metabolic disorders. In the absence of 27OHChol due to CYP27A1 deficiency, the human body manifests a disease known as Cerebrotendinous xanthomatosis, which is characterized by abnormal fat accumulation in the brain and joints [58]. Hence, excessive inhibition of sterol 27-hydroxylase to reduce 27OHChol levels is likely to result in secondary adverse effects. We suggest that it is imperative to develop drugs targeting the intermediate stages of 27OHChol-induced meta-inflammation to regulate chronic metabolic disorders.
 XML Download
XML Download