

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
The brain occupies approximately 20% of the whole-body blood supply and accounts for 20% of the basal metabolic rate.12 When the blood supply to the brain is compromised for approximately 5 minutes in normothermia, the brain experiences irreversible injuries. Thus, the application of hypothermia to the brain before or after cerebral ischemia has been attempted to reduce neurological injury and protect the central nervous system by lowering the body temperature as a means to lower the metabolic rate.
It is well known that the resting metabolic rate decreases to approximately 30–40% when the body temperature decreases to 33–34°C.3 In addition to this change in the overall metabolic rate, many researchers have tried to explore the alteration of specific metabolic pathways in hypothermia.14 However, most of these studies were in vitro or animal experiments, and several studies that were performed in humans were associated with ‘therapeutic hypothermia’ in patients with severe traumatic brain injury, in whom the target temperature ranged only from 33 to 35°C.5678 Thus, little information is known about the metabolic changes that occur in the human brain in conditions of hypothermia below 30°C.
Deep hypothermic circulatory arrest, along with selective antegrade cerebral perfusion, is the key strategy used during thoracic aortic surgery.91011 Surgery for the thoracic aorta might be the only situation in which in vivo studies evaluating metabolic changes in the human brain during deep hypothermia are feasible.
Therefore, this study was conducted to evaluate the changes in the metabolite profile in the human brain in deep hypothermia down to 25°C.
METHODS
Study population
This study was designed to enroll 13 patients considering the maximal number of patients who could be analyzed with 1 set of metabolite analysis kits. Patients over 19 years of age who were scheduled to undergo thoracic aortic surgery on a nonemergency basis and for whom selective antegrade cerebral perfusion under deep hypothermia was planned were screened for eligibility. From September 2019 to November 2020, overall 21 patients were screened for eligibility, 8 patients refused to participate, and 13 patients were enrolled in this study.
Operative strategy and sampling process
After anesthetic induction, median sternotomy was performed, and a right infraclavicular incision was also made to expose the right axillary artery. Axillary artery cannulation using a vascular graft was performed in all study patients. After arterial and venous cannulations, cardiopulmonary bypass and systemic cooling for hypothermia were initiated. Hemiarch or total arch replacement with selective antegrade cerebral perfusion under deep hypothermia (target temperature = 25°C) was performed in all study patients.
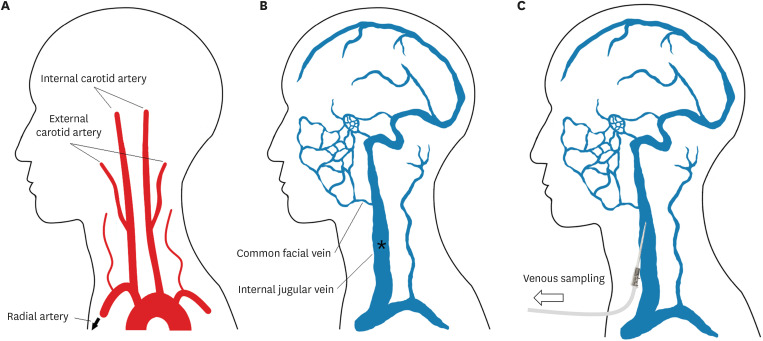
Arterial and venous blood was collected from the right radial artery and left internal jugular vein, respectively. A radial arterial catheter was inserted to monitor the arterial pressure during surgery, and an internal jugular venous catheter was inserted toward the cranial direction to closely sample the venous drainage from the brain. The metabolites supplied to the brain were assumed to be identical to those collected at the right radial artery, and the metabolites released from the brain were assumed to be identical to those collected at the left internal jugular vein (Fig. 1). Blood samplings were simultaneously performed to evaluate cerebral metabolite uptake or consumption by analyzing the differences between arterial blood supplied to the brain and venous blood released from the brain.12 Blood samples in normothermia were collected just after systemic heparinization (300 IU/kg) for axillary cannulation. Those in hypothermia of 30°C and 25°C were collected during systemic cooling. The blood samples for hypothermia were collected at 30°C and 25°C based on the nasopharyngeal probe which was considered to reflect the brain temperature, and circulatory arrest was performed at 25°C based on the urinary bladder probe which was considered to reflect the body temperature. Because the nasopharyngeal temperature drops more rapidly by systemic cooling than the bladder temperature does, all samples were obtained before the initiation of circulatory arrest. Two milliliters of each blood sample was placed in ethylenediaminetetraacetic acid-coated vacutainers (BD Vacutainer; Becton, Dickinson and Company, Franklin Lakes, NJ, USA). All samples were collected with minimal negative pressure and extreme caution to avoid any hemolysis.
Fig. 1
Illustration of the sample collection process. (A) The brain is mainly perfused by both internal carotid arteries and vertebral arteries, and the metabolites supplied to the brain were assumed to be identical to those collected at the right radial artery. (B) The metabolites released from the brain are mainly drained to the internal jugular vein, and the metabolites produced by the brain were assumed to be identical to those collected at the left internal jugular vein. (C) A short intravenous catheter for sampling from the internal jugular vein was placed as shown.
After sampling, the vacutainers were stored in a cold storage carrier and immediately transferred to the laboratory. Blood samples were immediately separated by centrifugation at 3,000 rpm for 10 minutes at 4°C, and the blood plasma components were collected. Centrifuged plasma samples were stored in −72°C liquefied nitrogen (N2).
Targeted metabolomics
A total of 188 metabolites were quantified using the AbsoluteIDQ®p180 kit (BIOCRATES Life Science AG, Innsbruck, Austria) in a combined liquid chromatographic mass spectrometry (amino acids and biogenic amines) and direct flow injection (acylcarnitines and glycerophospholipids) approach. The targeted metabolomics evaluated 21 amino acids, 21 biogenic amines, 40 acylcarnitines, 90 glycerophospholipids, 15 sphingomyelins, and 1 hexose. Detailed protocols for quantification complied with the manufacturer’s instructions. In brief, 10 μL of each sample was loaded onto a 96-well kit plate containing the internal standards. After evaporation and derivatization with the phenylisothiocyanate reagent, the metabolites were extracted using 5 mM ammonium acetate in methanol prior to analysis. Samples were then analyzed with a Waters® ACQUITY UPLC® I-Class system needle (Waters Corporation, Milford, MA, USA), composed of a binary solvent manager and a sample manager with flow-through, coupled with a SCIEX Triple Quad™ 5500 mass spectrometer (Danaher Corporation, Washington, D.C., USA).
Metabolomic data analysis
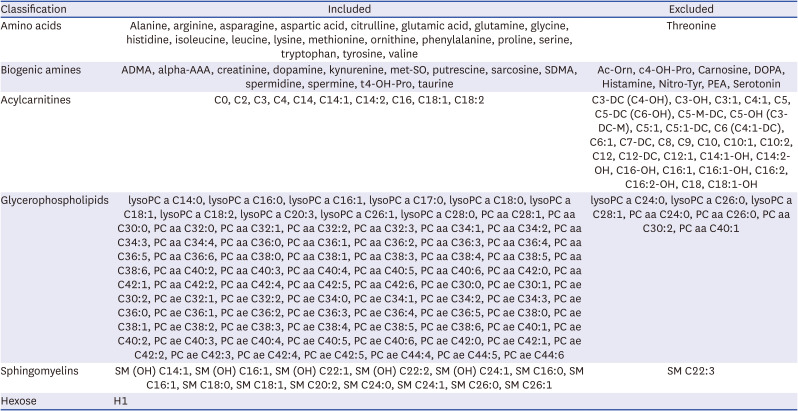
Analysis results were processed with Analyst® 1.6.3 Software from SCIEX and MetIDQ™ Oxygen from BIOCRATES (BIOCRATES Life Science AG). QC samples included in the kit were used to assess the quality control of the analysis and to further normalize the resulting data. The concentration data of each metabolite were then exported for manual filtering. Among 188 metabolites that were quantified in targeted metabolomics, 140 metabolites, including 20 amino acids, 13 biogenic amines, 10 acylcarnitines, 82 glycerophospholipids, 14 sphingomyelins and 1 hexose, were finally included in the analysis after excluding those with concentrations below the limit of detection in more than 80% of samples (Table 1). The ratios of metabolite concentrations in venous blood samples to those in arterial blood samples (V/A ratios) were calculated, and log2 transformation of the ratios [log2(V/A)] was performed for the comparison between the temperature groups. If the value of log2(V/A) for a metabolite was positive, it was interpreted as a release of the metabolite from the brain; if negative, it was interpreted as an uptake of the metabolite by the brain.
Table 1
List of metabolites evaluated in the targeted metabolomics performed in this study
Statistical analysis
Statistical analyses were performed using SPSS software (version 25.0; IBM, Armonk, NY, USA), MetaboAnalyst (version 5.0; a web-based software),13 and STATA (version 17.0; Stata Corp, College Station, TX, USA). The log2(V/A) values for each metabolite are presented as the mean ± standard deviation (SD). For clinical data, continuous variables are presented as the mean ± SD for normally distributed variables, and the median with interquartile range for nonnormally distributed variables. Categorical variables are presented as numbers and proportions.
RESULTS
Patient characteristics and operative data
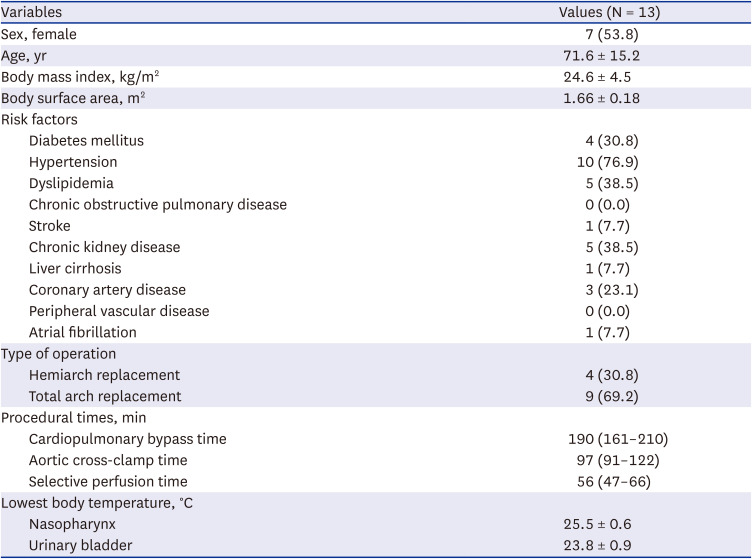
The mean age of the study patients was 71.6 ± 15.2 years and 7 patients (53.8%) were female. Comorbidities included hypertension (n = 10, 76.9%), dyslipidemia (n = 5, 38.5%), and chronic kidney disease (n = 5, 38.5%). The lowest body temperature during the surgery was 25.5 ± 0.6°C (Table 2).
Table 2
Preoperative characteristics, risk factors, and operative data of the study patients
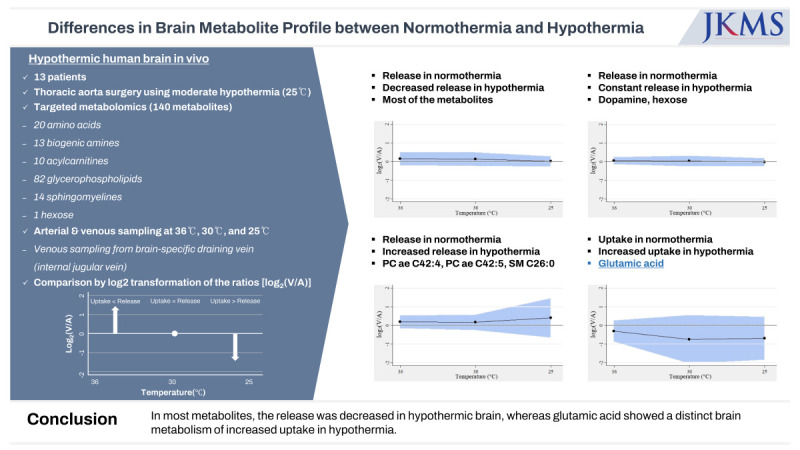
Changes in the metabolite profile during hypothermia
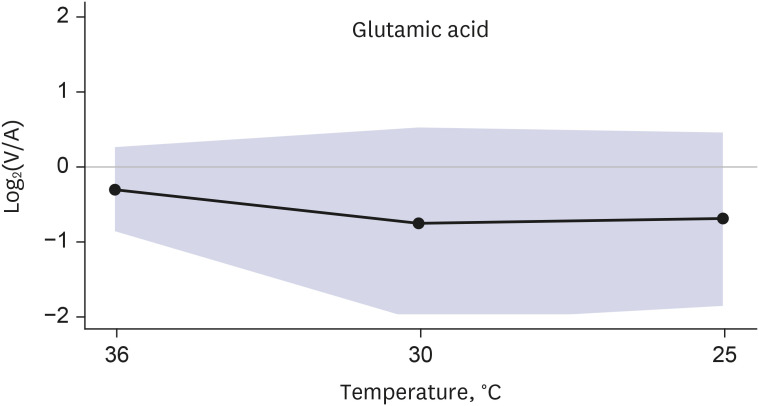
Of 140 metabolites analyzed, 137 metabolites were released from the brain in normothermia. Two glycerophospholipids (lysophosphatidylcholine a C14:0, phosphatidylcholine ae C30:0) were equivocal (defined as an absolute value of log2(V/A) < 0.05), and one amino acid (glutamic acid) was taken up by the brain in normothermia.
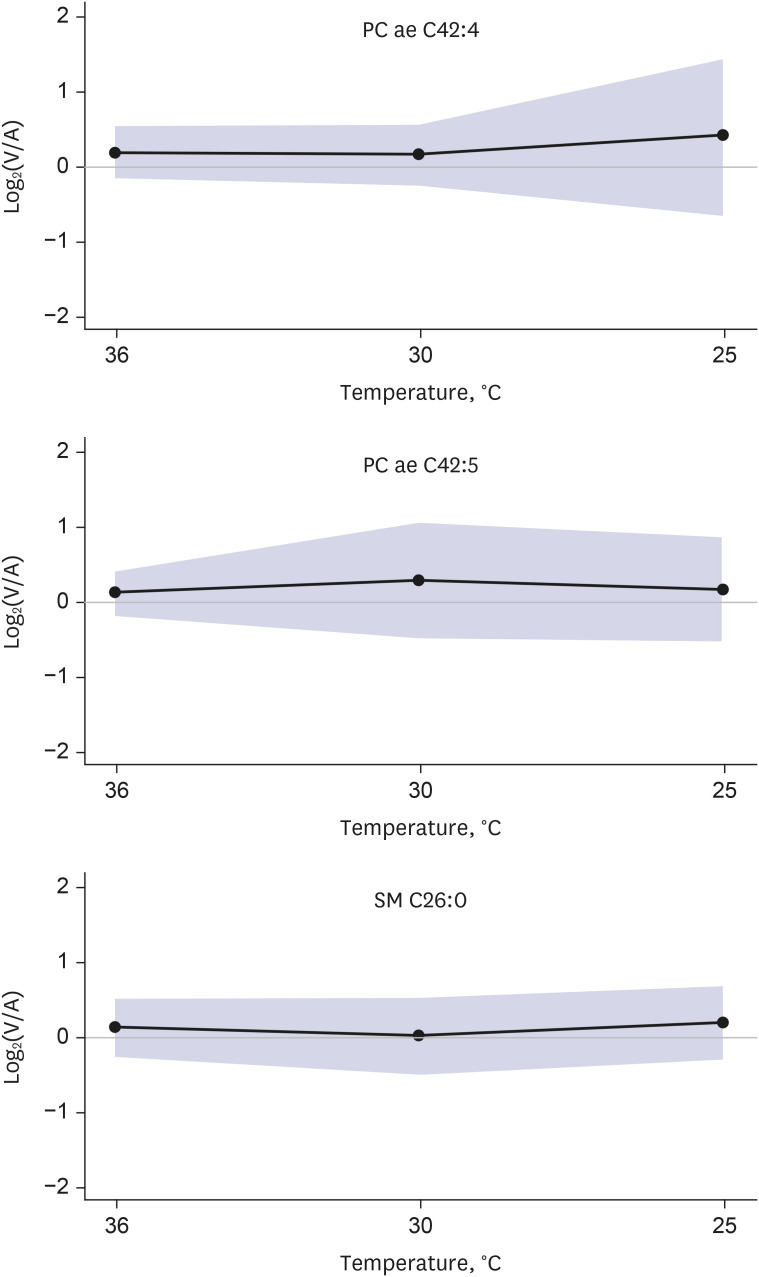
In the hypothermic state, the release of the metabolites was decreased for 132 out of 137 metabolites, whereas 2 metabolites, including 1 biogenic amine (dopamine) and 1 hexose, were constantly released from the brain (Fig. 2). For the other 3 metabolites, including 2 glycerophospholipids (phosphatidylcholine ae C42:4 and ae C42:5) and 1 sphingomyelin (C26:0) that were released from the brain in normothermia, an increase in release in the hypothermic state was observed (Fig. 3).
Fig. 2
Unlike most of the other metabolites that showed decreased release in hypothermia, the release of 2 metabolites (dopamine and hexose) from the brain was not decreased but was constant in hypothermia.
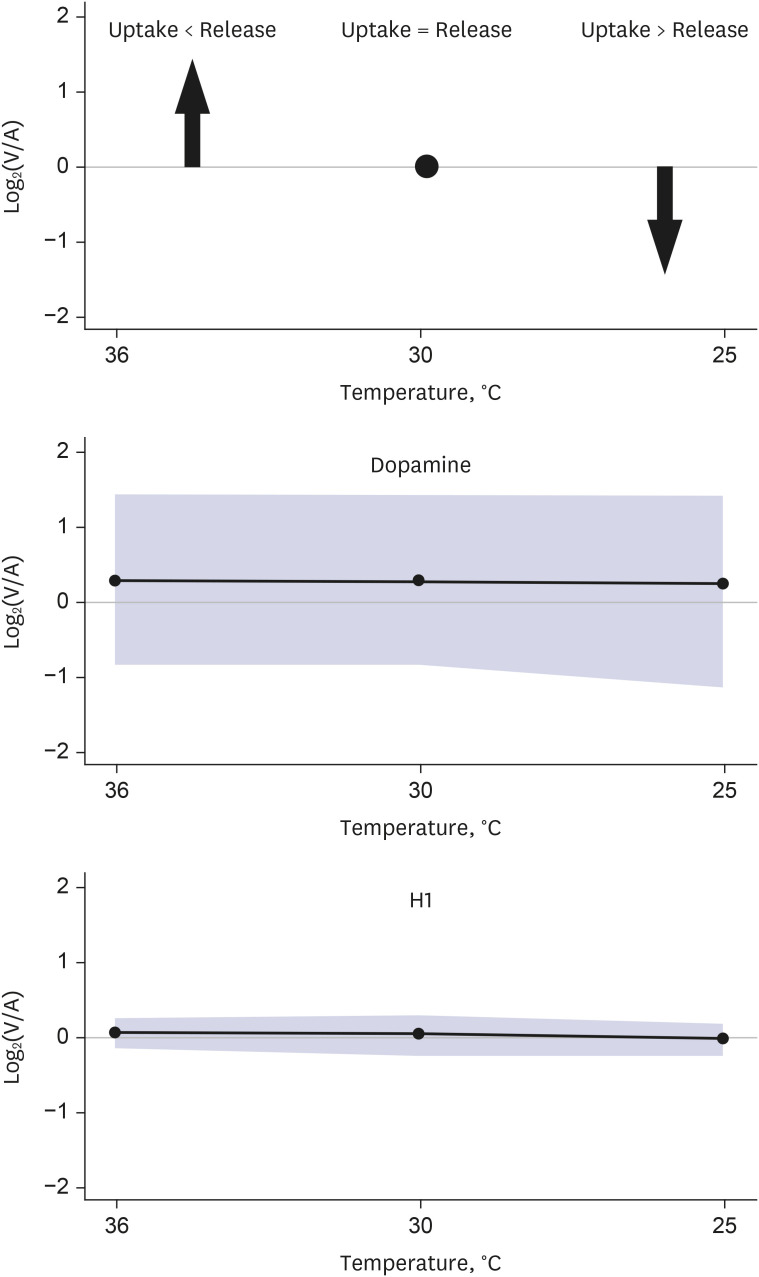
Fig. 3
Unlike most the of other metabolites that showed decreased release in hypothermia, the release of 3 metabolites (phosphatidylcholine ae C42:4, phosphatidylcholine ae C42:5, and sphingomyelin C26:0) from the brain changed to uptake by the brain in hypothermia.
Two glycerophospholipids (lysophosphatidylcholine a C14:0, phosphatidylcholine ae C30:0) that were equivocal in normothermia were also constantly equivocal during hypothermia. In one amino acid (glutamic acid) that was taken up by the brain in normothermia, the uptake increased during hypothermia (Fig. 4).
Clinical outcomes of the patients
There was no operative mortality. Three patients manifested with postoperative stroke; however, it was not likely that significant cerebral ischemia occurred during the operation. One patient suffered from subdural hemorrhage, another patient presented left hemianopia by acute cerebral infarction at postoperative day 3, and the other patient manifested with right side weakness at postoperative day 13 (Supplementary Table 7).
There were 2 patients who presented delirium immediately after the operation. When the change of glutamic acid concentration was reanalyzed after excluding these 2 patients, similar results were obtained: the log2(V/A) values for glutamic acid were −0.226, −0.266, and −0.287 at 36°C, 30°C, and 25°C, respectively.
DISCUSSION
The present study demonstrated 3 main findings. First, 137 out of 140 metabolites were released from the brain under normothermic conditions. Second, this release decreased in 132 of 137 metabolites as the body temperature decreased. Third, glutamic acid had a distinct cerebral metabolism; it was taken up by the brain in normothermia, and the uptake was intensified as hypothermia progressed (Fig. 5).
Fig. 5
To evaluate the difference in the brain metabolite profile between normothermia and hypothermia reaching 25°C, we performed targeted metabolomics of the hypothermic human brain in vivo. Thirteen patients who underwent thoracic aorta surgery under moderate hypothermia were prospectively enrolled, and 140 metabolites were analyzed for each patient. The study results demonstrated various degrees of changes in the release of metabolites by the hypothermic brain. In most metabolites, the release was decreased in hypothermia, whereas glutamic acid showed a distinct brain metabolism of increased uptake in hypothermia.
Unlike genes and proteins, whose functions are subject to epigenetic regulation and posttranslational modifications, respectively, metabolites serve as direct signatures of biochemical activity, and it is easy to correlate them with phenotype.14 Using a metabolomics approach, the aim of this study was to find changes in brain metabolism from a normothermic to a hypothermic status.15
Because the ultimate goal of lowering body temperature is to protect the brain by lowering cerebral metabolism, the characteristic changes in the brain metabolic pool could possibly provide valuable clues about the impact of hypothermia on metabolism and the mechanisms of neuroprotection or neurologic injury. Furthermore, if a set of characteristic biomarkers in the corresponding model is established, treatment effectiveness at the metabolic level could also be assessed.161718 For example, a previous study reported that nine characteristic metabolites, including five inhibitory neurotransmitters, two antioxidants, and two hormones, might represent biomarkers associated with brain protection.19
To estimate the metabolite exchanges in the brain, we performed metabolomics of blood samples obtained from the arterial circulation and brain-specific draining veins for the quantification of organ-specific metabolite production and consumption.12202122 The results of the present study showed the unique features of glutamic acid metabolism. The brain took up glutamic acid from the blood circulation, whereas it released almost all other metabolites.
Glutamic acid is mainly found in the cerebral cortex and dorsal spinal cord,23 and it plays multiple roles in the mammalian brain, including 1) as an excitatory neurotransmitter; 2) as the immediate precursor of the inhibitory neurotransmitter γ-aminobutyric acid; 3) as an essential component of intermediary metabolism; 4) as a building block of proteins; 5) as an energy substrate; and 6) paradoxically, as a potent neurotoxin.24 Among these multiple roles, glutamic acid is the main excitatory neurotransmitter, and as an intersynaptic message-transmitting substance synthesized in neurons, it is released into the synaptic gap via presynaptic membrane terminals and acts on receptors on the postsynaptic membrane to produce postsynaptic potentials.25 However, excessive extracellular glutamic acid results in excitotoxicity and neuronal apoptosis.26 It is well known that many brain diseases are associated with neurotransmitter abnormalities, mainly abnormal concentrations of glutamic acid.27
Previous studies demonstrated that the content of excitatory amino acid neurotransmitters, particularly glutamic acid, increased in the extracellular fluid during cerebral ischemia.12829 The large accumulation of glutamic acid leads to a continuous depolarization of neurons, causing a positive feedback effect that leads to a significant increase in the release of glutamic acid in neurons.23 A series of chain reactions, including impaired cellular energy metabolism, toxic effects of excitatory amino acids, inflammatory responses, calcium overload, free radical response, and ultimately cell death, are key mechanisms of the brain damage caused by increased levels of brain glutamic acid.30
A similar vicious cycle toward neuronal injury might occur in the hypothermic brain down to 25°C, in which we observed an increased uptake of glutamic acid to the brain in this study. The uptake of glutamic acid to the brain means that it is taken up from the intravascular space to extravascular space of the brain, which may mainly increase the extracellular level of the brain. Increased uptake of glutamic acid by the hypothermic brain might explain the clinical manifestation of minor or major neurologic symptoms, such as delirium, cognitive dysfunction, or stroke, after systemic hypothermia during cardiac surgery.
Following the aforementioned hypothesis, the suppression of glutamic acid uptake by the hypothermic brain might be helpful to prevent neuronal injury after hypothermia. If a pharmacologic agent reducing the extracellular concentration of glutamic acid was introduced and used in practice, it might help further brain protection in hypothermia. A previous study31 evaluated glial glutamate transporter-1 (GLT-1) which plays a major role in maintaining a low concentration of extracellular glutamic acid in the extracellular space, and the findings demonstrated that the upregulation of GLT-1 expression produced a neuroprotective effect by lowering extracellular glutamic acid levels in an animal experiment. In the same study, pretreatment with a functional antagonist of GLT-1 also resulted in partial reversal of the neuroprotective effect. Another study32 evaluated the expression of GLT-1 by astrocytes under hypothermic conditions in an in vitro model and demonstrated that the upregulation or downregulation GLT-1 expression in hypothermia was dynamic and dependent on the presence of neurons and neuronal activation. Transcriptional mechanisms that contribute to this process are still under investigation, and the findings of a previous study33 suggested that nuclear factor-κB might play a role in the neuron-dependent induction of GLT-1 expression in astrocytes. Therefore, to improve neurologic outcomes following hypothermia in the human brain, it will be important to identify the molecules and corresponding signal transduction pathways that affect astroglial GLT-1 expression in detail and to translate these findings into the development of new pharmaceuticals that are applicable in clinical practice.
In the perspective of clinical practice, there is no consensus on the ideal temperature for neuroprotection during aortic surgery although the benefit of hypothermia is widely accepted without doubt. Theoretically, cerebral metabolism increases linearly as the brain temperature rises, which causes greater consumption of oxygen, adenosine triphosphate, and glucose, leading to more ischemia.34 In contrast, drop in body temperature increases the risk of coagulopathy, hyperglycemia, acidemia, and total cardiopulmonary bypass time. Each surgeon usually adopts different temperature management methods according to the surgical strategies and the characteristics of individual patients.35
In the perspective of changes in metabolite profiles, little has been explored about the optimal temperature for the best neuroprotection. Although it is well-known that every drop of 1°C in body temperature reduces the ‘overall’ cerebral metabolic rate of oxygen by 6% to 7%,34 the behavior of each metabolite is still unexplored. In this study, this issue was addressed with targeted metabolomics and estimating the metabolite concentration at 3 different temperature points. However, it was not a continuous measurement down to 25°C, and also, there might be a possibility that a critical temperature exists beyond 25°C at which the extreme change of the metabolite profile occurs. Further investigation is required.
There are several limitations of the present study that should be noted. First, the sample size was relatively small. Second, the concentrations of respective metabolites in each individual might vary. Third, we did not analyze any association between metabolic changes and neurologic clinical outcomes. Fourth, the metabolites were collected from patients under the conditions of general anesthesia and cardiopulmonary bypass, which might affect our study results.
Targeted metabolomics demonstrated various degrees of changes in the release of metabolites by the hypothermic brain. For most metabolites, the release was decreased in hypothermia, whereas glutamic acid showed a distinct brain metabolism in that it was taken up by the brain in normothermia, and the uptake was increased in hypothermia.
 XML Download
XML Download