

 PDF
PDF Citation
Citation Print
Print


INTRODUCTION
The trigeminal subnucleus caudalis (Vc), also called medullary dorsal horn due to its homologous structure with the spinal dorsal horn, is known to be a major relay site for orofacial nociceptive inputs [1,2]. The sensory nucleus of the spinal cord, substantia gelatinosa (SG; lamina II), was named by Rolando for its translucent, gelatinous appearance [3]. SG neurons are commonly reported to receive incoming painful information and transmit it to ascending neurons [4]. In particular, SG neurons in Vc are known to play an important role in transmitting orofacial nociception to higher brain centers via thin myelinated Aδ and unmyelinated C primary afferent fibers [5].
Orofacial pain is frequently and severely triggered by the abundance of sensory receptors located in skin, mucous membranes, muscles, and ligaments of face [6,7]. For example, orofacial diseases such as trigeminal neuralgia, temporomandibular joint disorder, atypical toothache, and headache cause severe acute and chronic pain despite various treatments and reduce quality of life [7]. GABA is a major inhibitory neurotransmitter in the central nervous system (CNS) and is known to act as a nociceptive modulator in the spinal cord and human brain [8]. Additionally, many reported studies have shown that GABA can enhance or reduce pain transmission and perception due to differences in the composition of GABA receptor subunits [9,10].
The orofacial anti-nociceptive mechanisms of natural compounds related to neuromodulation have been studied over the past few decades [11], among which the prominent flavanone, (4',5,7-thrihydroxyflavanone (naringenin; Fig. 1) is majorly found in grapefruit and some other fruits and herbs, such as sour orange, tart cherries, tomatoes, cocoa, Greek oregano, and even beans [12,13]. Owing to its good bioavailability, naringenin is easily detected in human serum after intaking [14]. Interestingly, naringenin possesses anti-oxidative competence via protecting lipids against oxidative damage [15]. Furthermore, a previous study has shown that naringenin can advance memory function and reduce amyloid plaques and tau proteins in an Alzheimer’s disease model [16]. Naringenin also shows analgesic and anti-inflammatory activities through inhibiting pain-like behavior produced by inflammatory stimuli [17]. Although naringenin has been demonstrated to modulate nociceptive pain [18,19], role of naringenin in orofacial pain perception has not been reported yet. Therefore, the purpose of this study was to investigate the effect of naringenin and related mechanism on SG neurons of the Vc using the whole-cell patch-clamp technique and thereby gaining insight into its role in orofacial pain perception.
METHODS
Experimental animals and preparation of brainstem slices
Experiments were approved by the Institutional Animal Care and Use Committee of Jeonbuk National University (CBNU 2020-0122) and performed in accordance with the guidelines for the Care and Use of Laboratory Animals issued by Jeonbuk National University. Immature ICR mice postnatal days (PND) 7 to 21 were housed under 12-h light/dark cycles. They were allowed free access to food and water ad libitium (lights on at 7:00). Because the inhibitory effect in dorsolateral funiculus stimulation descends when spinal dorsal horn neurons closely approach to adult condition (from three postnatal weeks) [20,21], we referred the point of three weeks aging as a dividing point of maturity in SG neurons of mice.
Brain slices were prepared as described in our previous studies [22]. Mice were decapitated between 10:00 AM to 12:00 PM UTC + 9:00 (Universal Time Coordinate). Their brains were immediately excised and rapidly immersed in ice-cold artificial cerebrospinal fluid (ACSF) with the following compounds (in mM): 126 NaCl, 2.5 KCl, 2.4 CaCl2, 1.2 MgCl2, 11 D-glucose, 1.4 NaH2PO4, and 25 NaHCO3 (pH 7.3–7.4, bubbled with 95% O2 and 5% CO2). To prepare coronal slices (180–200 µm in thickness), brainstem containing the rostral part of Vc was glued to an agar block and then, cut in ice-cold ACSF with a vibratome (Leica VT1200S; Leica Biosystems). Before electrophysiological recordings, these slices were allowed to recover in oxygenated ACSF for a minimum of one hour. All experimental steps were performed at room temperature.
Electrophysiology recoding
Brain slices were transferred to a recording chamber, which was continuously superfused with oxygenated ACSF at 4–5 ml/min. Slices were viewed on the stage of an upright microscope (BX51W1; Olympus) with differential interference contrast optics. Patch pipettes were pulled from borosilicate glass capillaries (PG52151-4; WPI) using a Flaming/Brown micropipette puller (P-97; Sutter Instruments Co.). A high chloride-based internal solution, consisting of 140 mM KCl, 1 mM CaCl2, 1 mM MgCl2, 10 mM HEPES, 4 mM Mg-ATP, and 10 EGTA (pH 7.3 with KOH) was filtered through a disposable 0.22 µm filter and loaded into the pipette.
Under the microscope’s magnification, the SG (lamina II) was apparently distinguished as a translucent band, just medial to the spinal trigeminal tract crossing the lateral edge of the brain slice. Tip resistance of recording pipettes ranged from 4 to 6 MΩ when loaded with a high chloride-based internal solution. Once a gigaseal was achieved between SG neuron and the pipette, neurons were voltage-clamped at –60 mV and current-clamped for whole-cell patch-clamp recording. An intact neuronal membrane was ruptured by a short suction for whole-cell patch-clamp recordings [23].
Signals were amplified and filtered at 1 kHz with an Axopatch 200B (Molecular Devices) and digitized using a Digidata 1440A (Molecular Devices). Acquisition and subsequent analysis of the recorded data were done using Clampex 10.6 software (Molecular Devices). The validity of this process was confirmed by once more visual analysis of all traces before being approved for further analysis in Origin software (OriginLab Corp.).
Reagents
Naringenin, GABA and chemicals for ACSF were purchased from Sigma Aldrich. Stocked chemicals were prepared according to their solubility in dimethyl sulfoxide (DMSO) or distilled water and diluted to tested concentration in ACSF shortly before being bath applied.
Data analysis and statistics
To determine relative changes, the target response was divided by its corresponding control response. SG neurons were considered affected if the presence of naringenin resulted in a change of more than 10% compared to the control response. For area under the curve (AUC) measurement, the negative pA value was converted to a positive value, and then the mean value of 1-min period before GABA application was established as the baseline mean value. Traces recovered within 6 min from the time of GABA in presence of naringenin were utilized for AUC calculation. Each data point was normalized to the baseline mean value and Prism 10's AUC calculation method was employed to determine the AUC.
The means of two groups were compared using either paired or unpaired Student’s t-test, and for comparisons involving multiple groups, a one-way ANOVA followed by the post-hoc Scheffe test was applied. All numerical values are expressed as the mean ± standard error of the mean (SEM). Statistical significance was defined as p < 0.05 and the levels of significance were defined by asterisks (*p < 0.05, **p < 0.01, and ***p < 0.005).
RESULTS
Effect of naringenin and GABA in the SG neurons of the Vc
To confirm the effect of naringenin, various concentrations (ranging from 1 µM to 2 mM) were bath applied to SG neurons in voltage clamp mode. Bath application of naringenin induced weak inward current with the concentration 1 mM and greater (Fig. 2A, B, n = 10). Subsequently, the effect of 0.5 mM naringenin on holding current was assessed, revealing no significant impact (Fig. 2C, D, n = 3). DMSO, used as a vehicle for naringenin, also demonstrated no effect on membrane current (figure not shown). Next, bath application of 30 µM GABA for 3 min in SG neurons induced inward currents of two distinct characteristics, i.e., slow response (Fig. 2E) and fast response (Fig. 2F). The average time for the slow GABA response was 112.2 ± 4.96 sec (n = 15), while it was 40.2 ± 4.08 sec (n = 6) for fast response. The mean amplitude of slow GABA response was –52.6 ± 9.84 pA (n = 15), and the fast GABA response had a mean amplitude of –53.7 ± 18.06 pA (n = 6). However, repeated application of both slow (Fig. 2G, n = 15) and fast (Fig. 2H, n = 6) GABA-induced response showed no significant differences (total, Fig. 2I; n = 21, p > 0.05). The results in Fig. 2E–I suggest that the response of increasing or decreasing in the subsequent process is attributable to the concurrent application of naringenin with GABA, as there is no significant change observed with repeated application.
Effect of naringenin on GABA response in the SG neurons
To investigate the effect of naringenin on GABA response, GABA was first bath applied and washed out for 5 min. Next, naringenin 0.5 mM was bath applied for 3 min and then coapplied with GABA. As shown in Fig. 3, 30 µM GABA-induced slow response was increased in 57% of neurons tested (Fig. 3A, n = 12/21). In this case, the mean relative amplitude and AUC were significantly increased to 1.70 ± 0.185 (Fig. 3B; n = 12, p < 0.01) and 1.88 ± 0.265 (Fig. 3C; n = 12, p < 0.01), respectively in the presence of naringenin. Next, naringenin decreased the GABA-induced fast response in 33% of SG neurons tested (Fig. 3D, n = 7/21). In this case, the mean relative amplitude and AUC were significantly decreased to 0.589 ± 0.050 (Fig. 3E; n = 7, p < 0.001) and 0.757 ± 0.075 (Fig. 3F; n = 7, p < 0.05), respectively in the presence of naringenin. In two neurons, no significant change was observed even in the presence of naringenin (n = 2/21, 10%, data not shown).
Concentration dependent effect of naringenin on GABA response on the SG neurons
As observed in Fig. 3, naringenin exhibited varied effects on GABA induced inward current depending upon the nature of GABA response. Fast GABA responses were suppressed while slow GABA responses were enhanced by 0.5 mM naringenin. Next concentration dependent effect of naringenin was assessed in both form of GABA response. Naringenin concentration ranging from 1 µM to 1 mM exhibited concentration dependent effect on both slow and fast GABA responses. Slow GABA response was enhanced (Fig. 4A) while fast GABA response was suppressed (Fig. 4B) with increasing concentration of naringenin. Fig. 4C and D represent the histograms showing the increasing and decreasing tendency of slow GABA response and fast GABA response respectively, with the increasing concentration of naringenin. The relative amplitude of GABA-induced inward currents exhibited distinct patterns in response to naringenin concentration. Specially, for slow GABA responses, the relative amplitudes were, naringenin 1 µM: 1.39 ± 1.06; 10 µM: 2.17 ± 0.950; 100 µM: 2.83 ± 1.55; 1 mM: 2.87 ± 1.09 (Fig. 4C, n = 4, p < 0.05; one-way ANOVA). In contrast, for fast GABA responses, the relative amplitudes demonstrated a decreasing trend with increasing naringenin concentration, naringenin 1 µM: 1.00 ± 0.088; 10 µM: 0.96 ± 0.244; 100 µM: 0.74 ± 0.221; 1 mM: 0.23 ± 0.086 (Fig. 4D, n = 5, p < 0.01; one-way ANOVA).
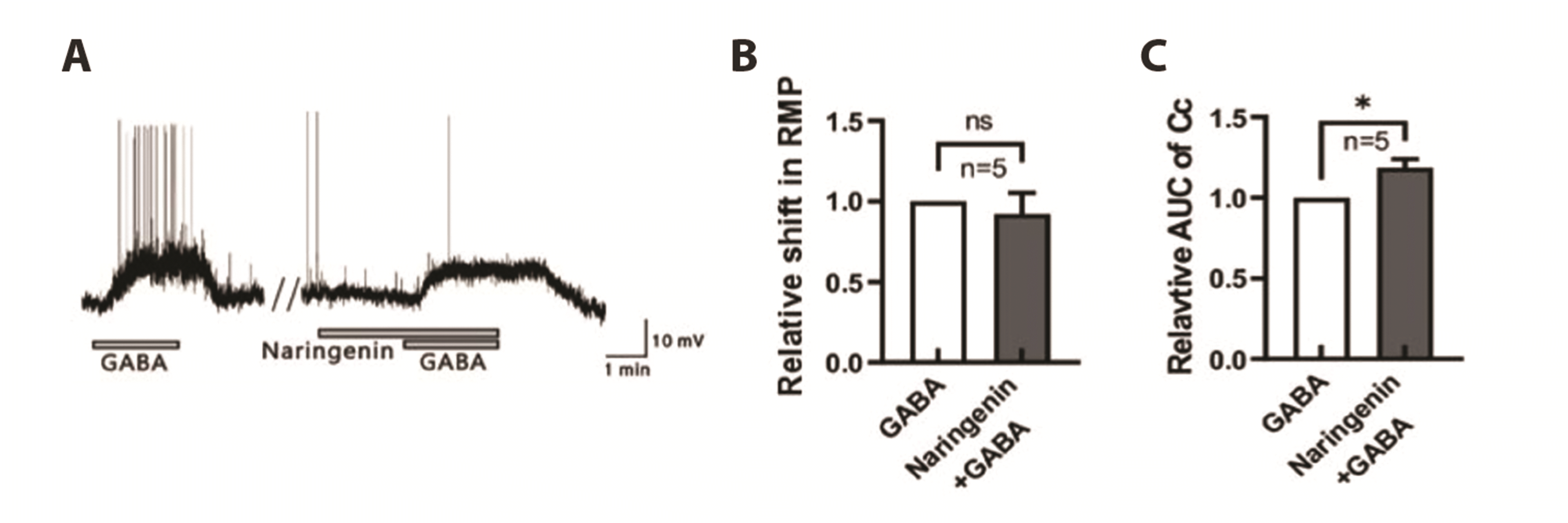
Enhance of durability the GABA receptor activity by naringenin on the SG neurons
To confirm that naringenin could affect membrane potential, GABA was applied in the presence of naringenin in current-clamp mode (Fig. 5). Naringenin itself did not influence membrane potential but enhanced the durability of GABA response as shown in Fig. 5A. Initially, GABA was bath-applied for 2-min, followed by a 5-min washout period, and then 0.5 mM naringenin was applied for 2 min before co-application with GABA. The GABA-induced depolarization did not exhibit a significant change, with a mean relative amplitude of 0.92 ± 0.130 (Fig. 5B, n = 5, p > 0.05). However, the mean relative AUC significantly increased to 1.23 ± 0.039 in the presence of naringenin (Fig. 5C, n = 5, p < 0.05). This result suggests that naringenin may have an enhancing effect on the inhibitory GABA receptors known to modulate SG neurons of the Vc.
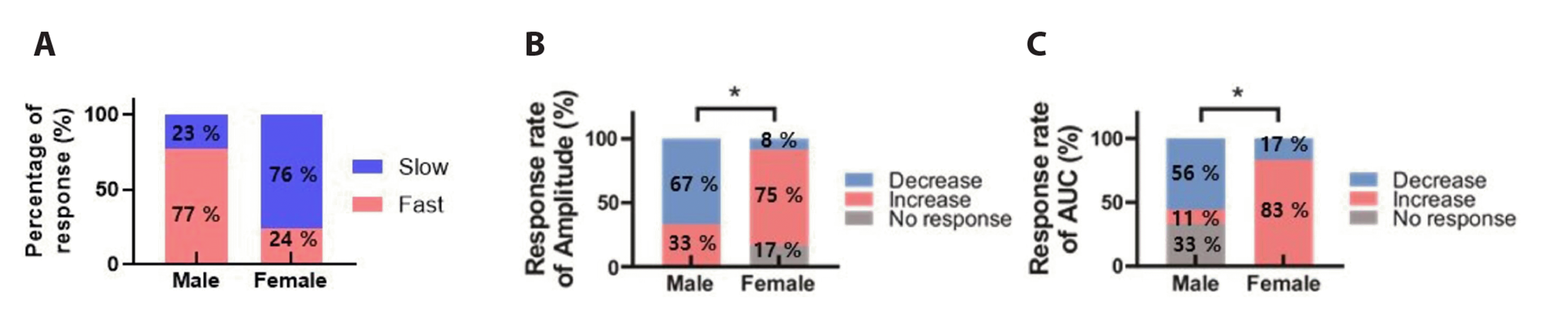
Naringenin exhibits sex-dependent effect on GABA mediated responses on the SG neurons
As shown in Fig. 6A, further analysis of the voltage-clamped recorded data revealed that most SG neurons from males (10/13, 77%) showed fast GABA response and remaining neurons (3/13, 23%) showed slow response. In contrast, most SG neurons (13/17, 76%) from female juvenile mice showed a slow GABA response upon bath application of 30 µM GABA, while the remaining neurons (4/17, 24%) displayed a fast GABA response.
As previously indicated in the results, naringenin inhibits fast GABA responses and enhances slow GABA responses. Among males, 67% of the tested SG neurons (6/9) exhibited a decrease in GABA response amplitude upon exposure to naringenin, while the remaining three neurons showed an increase. Conversely, in females, the application of 0.5 mM naringenin led to an increased amplitude of the GABA response in 75% of the tested SG neurons (9/12), with one neuron displaying a decrease, and no effect observed in the remaining two neurons (Fig. 6B). Additionally, naringenin predominantly reduced the AUC of GABA-induced inward currents in most of the tested neurons (56%, 5/9), with one neuron showing an increase, and the remaining three neurons remaining unaffected in males. In contrast, most SG neurons (83%, 10/12) from females demonstrated an increase in the AUC of the GABA-induced response in the presence of 0.5 mM naringenin, while the remaining two neurons displayed a decrease (Fig. 6C). This suggests that naringenin’s impact on SG neurons is influenced by the nature of the GABA response and exhibits a sex-dependent pattern.
DISCUSSION
In the present study, we observed that naringenin exerted an influence on GABA receptor-mediated responses within the SG neurons in the Vc. Notably, naringenin potentiated GABA receptor-mediated responses in the majority of SG neurons showing slower GABA responses, while it inhibited such responses in a minority of neurons exhibiting faster GABA response. Importantly, both the potentiation and inhibition of GABA receptor-mediated responses were found to be dependent upon the concentration of naringenin. Furthermore, our results underscore the sex-dependent nature of naringenin’s effect on GABA responses. These findings highlight the significant influences of the nature of the GABA response itself in determining the effects of naringenin on it.
This study focuses on SG neurons in the dorsal horn (lamina II) of the spinal cord, recognized as the central gate for modulating nociception information [24,25]. Laminae I-III of the dorsal horn contains numerous inhibitory interneurons that regulate sensory information before transmitting it to other parts of the brain and spinal cord. Most neurons in layers I-III are glycine-rich, and it is reported that these inhibitory interneurons also contain GABA. These interneurons constitute approximately 30% of laminae I-II neurons and about 40% of laminae III neurons. Immunocytochemical studies confirm their GABAergic nature, with some using glycine as a co-transmitter [24,26]. Several reports indicate that the development of the nervous system can vary based on sex, leading to diverse nerve structures [27]. Additionally, changes in GABA receptor responsiveness may occur depending on sex or age [28,29]. Chudomel et al. [28], observed higher GABAA receptor-mediated sIPSC amplitude and frequency in male rats compared to females in the substantia nigra reticulate, with rise and decay time decreasing with age. This variation according to sex and age may depend on the heterogeneity on the GABAA receptor subunit composition [22].
GABA receptors are pentameric protein complexes composed of five subunits assembled from the pool of 19 different subunits (α1–6, β1–3, γ1–3, δ, ε, θ, π and ρ1–3) [30,31]. The typical GABA receptors consists of five subunits, with general subunit stoichiometry of 2α, 2β, and 1γ [32-34], arranged pseudo-symmetrically around central ion pore [35]. Bohlhalter et al. [36] discovered that within the spinal cord, the GABAA receptor subunits α3, β2,3 and γ2 exhibited widespread expression, contrasting with the more confined lamina-specific distribution of the other three α subunits. Specially, α1 and α5 subunits were predominantly found in the intermediate region, while α2 subunits prevailed in the superficial layers of the dorsal horn and in somatosensory and preganglionic motoneurons. The unique expression patterns of GABAA receptor subtypes across laminar layers in the spinal cord suggests that the diversity of GABAA receptors plays a role in regulating nociception, sensory input, and motor control at the segmental level [36].
Several compounds have been identified and are claimed to modulate selectively individual GABA receptor subtypes. These compounds can modulate GABA-induced currents or directly activate GABA receptors by binding to allosteric sites on GABA receptors or directly interacting with the GABA binding site, respectively [37]. Benzodiazepines, anesthetics, neurosteroids, picrotoxin, and many other molecules can allosterically modulate GABA receptors [38]. Numerous studies have highlighted the possible effects of flavonoids on the CNS via modulation of GABA receptor activity [8,39]. For example, flavonoids such as apigenin, 6-methylapigenin, dinatin, skrofulein, cirsilineol, hispidulin, and naringenin show affinity for GABA-benzodiazepines sites [11,40-42]. The 2ʹ-hydroxyl group of flavonoids is reported as relevant for its benzodiazepine binding affinity. Wang et al. [43] suggested that flavonoid baicalin-induced responses on SG neurons were mediated via the activation of the benzodiazepines site of GABA receptors [43,44]. However, some studies also report that flavonoids such as quercetin and apigenin affinity for the GABA receptor is independent of the benzodiazepine site [45,46]. Such benzodiazepine-independent mechanism is reported in responses mediated by α1β1γ2s GABA on Xenopus oocytes [45] and α1β2γ2 GABA on HEK293 cells [46].
Naringenin, a citrus flavonoid, exhibits various effects on the CNS and can bind to GABA receptors, exerting anxiolytic effects due to its ability to permeate the blood-brain barrier [47-49]. In this study, we found that naringenin enhanced GABA-receptor-mediated currents and AUC in the majority of neurons tested. This enhancement of GABA receptor activities can be explained by naringenin’s allosteric modulation of GABA receptors as flavonoids are known to interact with benzodiazepine binding sites on GABA receptors [8]. In addition, radioligand binding assay reveals that substances like (S) naringenin, glabrol, and 8-lavandulyl- flavanones act on the GABA receptors’ benzodiazepine site [50]. Besides flavonoid naringenin, essential oil linalool [51], terpene borneol [52], and biphenyl honokiol [53] possess the potential to influence the GABA receptor-mediated action on SG neurons of Vc. Notably, 6-hydroxyflavone, a flavone, shows diverse responses towards GABA, acting as a subtype-selective partial positive allosteric modulator at the flumazenil-sensitive benzodiazepine site and flumazenil-sensitive partial inverse agonist or negative modulator at α1β3γ2, α2β3γ2, and α5β3γ2 but not at α3β3γ2 receptor subunits [8]. The sensitivity and response to GABA can vary depending on subunit combinations, as seen with antiepileptic drugs like carbamazepine and phenytoin, which modulate GABAARs in a subunit-dependent manner. Both carbamazepine and phenytoin enhance GABA-induced currents in cultured rat cortical neurons and HEK293 cells transiently expressing the GABAAR α1β2γ2 subtype, however; GABAAR α1β2 subtype remained unaffected, and minimal effects was observed for the α3β2γ2 and α5β2γ3 receptor isoforms [54].
Interestingly, naringenin showed varied effects on GABA response depending on sex. For most female SG neurons, naringenin only enhanced GABA responses, whereas both enhancing and inhibiting effects were observed in males. Benzodiazepines, GABA receptor allosteric modulators, binding pocket have been identified in the cleft between the α1 and γ2 subunits of the GABA receptors [55]. Ravizza et al. [56] reported higher expression of α1 subunit mRNA in substantia nigra pars reticulate regions of females compared to the male of the same age groups (PND 15 and PND 30). Based on this evidence, we may speculate that naringenin exhibited varied effects on GABA receptor action across sex might be dependent on GABA receptor subunits composition. However, no findings are reported for the sex-dependent variation in GABA receptor subunits in the SG region and require further research.
The results of this study imply that naringenin’s responsiveness to the GABA receptor differs by sex, and these findings suggest that pain control mechanisms may differ depending on sex. This variation might be dependent on the level of expression of GABA receptor subunits. The GABA receptors' pharmacology, pattern of expression in the brain, and biophysical properties including their affinity for GABA are all influenced by the subunit composition [57]. To conclude, considering that SG neurons of the Vc as a principal region for modulation of orofacial nociception, the sex-dependent effect of naringenin on GABA receptors might contribute to sex-dependent orofacial pain modulation, at least in part.

 XML Download
XML Download