

 PDF
PDF Citation
Citation Print
Print


INTRODUCTION
Diabetic nephropathy (DN) is the primary cause of chronic kidney disease (CKD) and seriously affects the prognosis and survival of patients with diabetes [1,2]. The incidence of DN may involve hemodynamic and metabolic factors that significantly increase the levels of blood glucose and advanced glycation end products (AGEs), consequently leading to the activation of the renin–angiotensin–aldosterone system [3]. Currently, the management of diabetes focuses on reducing proteinuria through maintaining blood pressure and blood glucose levels; however, these interventions are unable to prevent the development of CKD [4]. Therefore, developing safe and effective methods to manage DN is necessary.
Results of previous studies have indicated that mesangial cells (MMCs) play a vital role in controlling mesangial matrix homeostasis, glomerular filtration rate, and normal glomerular function through regulating extracellular matrix (ECM) and producing cytokines. The progression of DN is characterized by overproliferation of MMCs, accumulation of ECM, thickening of the basement membrane, and expansion of the mesangial area, leading to the development of glomerulosclerosis [5]. Early glomerular mesangial lesions, characterized by MMC proliferation, are among the most prominent pathological changes in DN [6].
Inflammation is stimulated in response to detrimental conditions to maintain tissue homeostasis and integrity [7]. Long-term chronic inflammation can lead to the progression of different diseases [8]. Accumulating evidence suggests that inflammation is involved in the progression of DN [9]. As reported previously, DN was accompanied with an increased production of pro-inflammatory cytokines, including tumor necrosis factor-α (TNF-α), interleukin (IL)-1β, and IL-6 [10]. Hence, inhibition of inflammation has become a potential strategy for the treatment of DN.
In recent years, various traditional Chinese medicines have been proposed to participate in multiple cellular activities such as cell proliferation, apoptosis, and differentiation [11]. Cornuside is a secoiridoid glucoside extracted from the fruit of Cornus officinalis, a traditional oriental medicine [12]. Crude extracts of C. officinalis have been shown to exert antineoplastic, anti-inflammatory, hepatoprotective, and anti-sepsis effects [13]. Of note, the results of a previous study revealed that loganin and its derivatives are the active compounds in the C. officinalis fruit and may exert anti-DN functions [14]. However, the role of cornusides in DN remains unclear.
In this study, we established an in vitro model of DN through treating MMCs with glucose and aimed to explore the possible underlying mechanism of the antidiabetic effects of cornuside.
METHODS
Cell culture
Mice MMC line SV40-MES13 was purchased from Procell and cultured in DMEM/F12 medium (Thermo Fisher Scientific) containing 10% fetal bovine serum and 1% penicillin–streptomycin at 37°C with 5% CO2. Cells were digested with trypsin for passaging when they reached approximately 90% confluence.
In vitro model of DN induced by glucose
MMCs in the logarithmic phase were treated with glucose (0, 10, 25, 50, and 100 mM) for 24 h to establish an in vitro DN cell model. Cells were cultured at 37°C with 5% CO2 for 24 h, followed by the subsequent analyses.
Cell counting kit-8 (CCK-8)
MMCs (5 × 103 cells/well) were cultured for 24 h under the different treatments. CCK-8 reagents (10 μl; Beyotime) were added into each well and maintained for 2 h. The absorbance was measured at 450 nm using a microplate reader (Bio-Rad).
5-Ethynyl-2′-deoxyuridine (EdU)
MMCs were cultured in 96-well plates and treated with 50 µM of EdU-labeling medium for 2 h. After treatment with 4% paraformaldehyde and 0.5% Triton X-100, the cells were stained with an anti-EdU working solution and photographed using a fluorescent microscope (Olympus).
Enzyme-linked immunosorbent assays (ELISA)
The expression levels of IL-6, IL-1β, and TNF-α were examined using commercially available assay kits (Thermo Fisher Scientific). Briefly, cell culture supernatants were collected and added to a 96-well plate coated with IL-6, IL-1β, and TNF-α antibodies for incubation and then washed with PBS containing 0.1% Tween 20. After reacting with primary and secondary antibodies, a chemiluminescent substrate was added and incubated at 37°C for 15–30 min. The optical density was measured using a microplate reader (Bio-tek) at 450 nm after the addition of the stop solution.
Reverse transcription quantitative real-time polymerase chain reaction (RT-qPCR)
Trizol reagent (Invitrogen) was used to extract total RNAs. The RNA samples were then reverse transcribed into cDNA, followed by quantitative PCR using a commercially available kit (One Step TB Green PrimeScript RT-PCR Kit [Perfect Real Time]; Takara). The sequences of the primers were: p21: F 5ʹ-TAAGGACGTCCCACTTTGCC-3ʹ, R 5ʹ-CGTCT CCGTGACGAAGTCAA-3ʹ; proliferating cell nuclear antigen (PCNA): F 5ʹ-GAACCTCACCAGCATGTCCA-3ʹ, R 5ʹ-ATTCACCCGACGGCATCTTT-3ʹ; cyclin D1: F 5ʹ-AAAATGCCAGAGGCGGATGA-3ʹ, R 5ʹ-GAGGGG GTCCTTGTTTAGCC-3ʹ; AKT: F 5ʹ-CCGCCTGATCAA GTTCTCCT-3ʹ, R 5ʹ-GCCACATGTGTGGTCTCAAAC-3ʹ; GAPDH: F 5ʹ-CCCTTA AGAGGGATGCTGCC-3ʹ, R 5ʹ-TTCCCATTCTCGGCCTTGAC-3ʹ.
Western blot
MMCs were lysed using RIPA lysates (Beyotime Biotechnology) and then separated by 10% SDS-PAGE, followed by transfer to PVDF membranes (Millipore). The membranes were sealed and then incubated overnight with the following primary antibodies, including PCNA (ab18197, Abcam), cyclin D1 (ab16663, Abcam), p21 (ab109520, Abcam), p65 (ab32536, Abcam), AKT (ab283852, Abcam), p-AKT (ab38449, Abcam), p-IκB-α (FS-K0390, affandi-e), COL1 (ab275746, Abcam), fibronectin (ab2413, Abcam), lamin A/C (ab108595, Abcam), and β-actin (ab8226, Abcam). Next, the membranes were washed with corresponding secondary antibodies and visualized using an ECL detection kit (Millipore).
RESULTS
Glucose treatment increased viability of MMCs
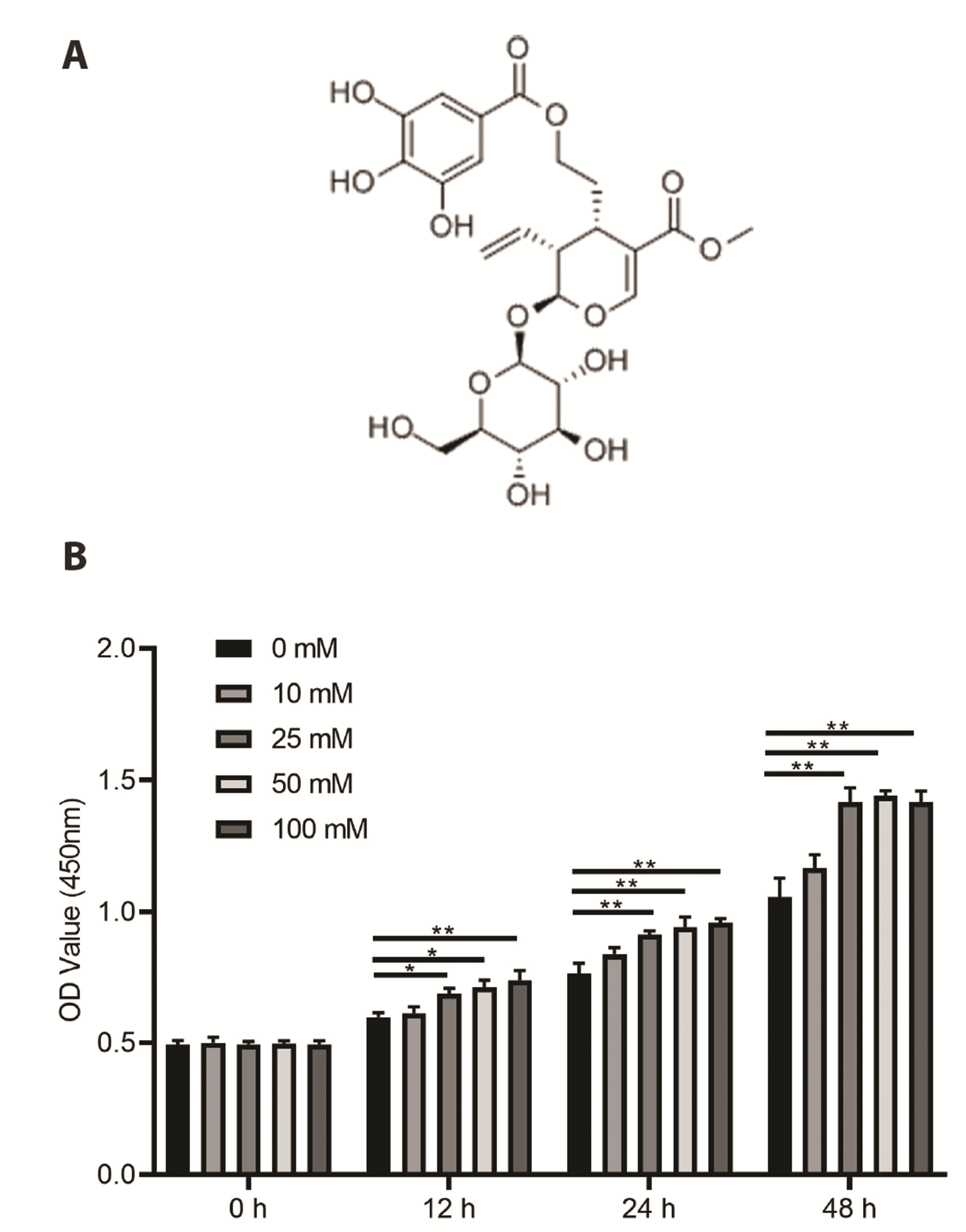
Fig. 1A shows the chemical structure of cornusides. Next, MMCs were treated with glucose (0, 10, 25, 50, and 100 mM) for 12, 24, and 48 h to establish an in vitro model of DN. We then assessed cell viability upon glucose treatment. We found that 25, 50, and 100 mM markedly increased the viability of MMCs in a time-dependent manner (Fig. 1B); however, the effects of 25, 50, and 100 mM were not significant at each time point. Therefore, 25 mM glucose was used in subsequent experiments.
Cornuside the chemical structure of cornuside high glucose-treated overproliferation of MMCs
Next, to evaluate the impacts of cornuside on the growth of glucose-treated MMCs, 25 mM of treated MMCs were co-treated with cornuside (5, 10 or 30 μM), and the viability as well as proliferation of the MMCs were determined using CCK-8 and EdU assays. We found that 25 mM glucose markedly increased the viability and proliferation of MMCs, whereas cornuside inhibited the viability (Fig. 2A) and proliferation (Fig. 2B) of glucose-treated MMCs in a dose-dependent manner.
Cornuside inhibited the production of IL-6, TNF-α, and IL-1β in high glucose-treated MMCs
We further evaluated the influence of cornuside on the expression of proinflammatory cytokines in high glucose-treated MMCs. As revealed in Fig. 3, high glucose stimulation significantly increased the expressions of IL-6, TNF-α, and IL-1β in contrast to the control group. However, cornuside significantly decreased the expression of IL-6, TNF-α, and IL-1β in glucose-treated MMCs in a dose-dependent manner.
Cornuside inhibited the AKT and NF-κB signaling pathways in high glucose-treated MMCs
We further examined the influence of cornuside on the activation of AKT and NF-κB signaling pathways in high glucose-treated-induced MMCs. Through RT-qPCR analysis, we observed that high glucose treatment significantly increased the mRNA expression of PCNA and cyclin D1 and suppressed p21 mRNA expression. Concurrently, co-treatment with cornuside markedly decreased the mRNA levels of PCNA and cyclin D1 and increased the mRNA level of p21 in a dose-dependent manner. Cornuside did not affect AKT mRNA expression (Fig. 4A). Moreover, high glucose increased the protein expression of PCNA, cyclin D1, p-AKT p-IκB-α, COL1, and fibronectin, while reducing protein expression of p21. Conversely, cornuside treatment decreased the protein expression of PCNA, cyclin D1, p-AKT, p-IκB-α, COL1, and fibronectin but enhanced protein expression of p21 in a dose-dependent manner (Fig. 4B). Additionally, results of western blot analysis also reflected the translocation of p65 into the nucleus upon high glucose induction (Fig. 4C). Concurrently, cornuside treatment obviously repressed the translocation of p65 in a dose-dependent manner.
DISCUSSION
Overproliferation of glomerular MMCs and inflammatory conditions are prominent pathological characteristics of DN [15]. High glucose can activate inflammatory pathways, e.g., NF-κB signaling pathway along with the increased production of pro-inflammatory cytokines [16]. C. officinalis is a dietary anti-inflammatory agent widely used clinically [17]. Previous studies have suggested that the fruit of C. officinalis can be used to treat backache, hypertension, and polyuria [18]. In addition, a previous study revealed the biological properties of C. officinalis, including anti-inflammatory, renal, and hepatic protective roles, and immunomodulation [19]. Studies have shown that Cornuside I can improve osteoporosis through inducing bone mesenchymal stem cell proliferation [20]. In this study, we established an in vitro model of DN, and the results of CCK-8 and EdU analyses showed markedly reduced glucose-induced proliferation of MMCs. These results suggest the anti-proliferative functions of cornuside.
Conversely, the immunomodulatory and anti-inflammatory activities of cornuside in various diseases have been discussed previously [21]. Kang et al. [22] reported that cornuside inhibited cytokine-induced pro-inflammatory effects in human umbilical vein endothelial cells. Zhang et al. [23] found that cornuside suppressed experimental autoimmune encephalomyelitis in the central nervous system. Moreover, Choi et al. [24] suggested that cornuside suppresses lipopolysaccharide-induced increases in inflammatory mediator production. In this study, we found that cornuside could alleviate high glucose-induced production of pro-inflammatory cytokines, i.e., IL-6, TNF-α, and IL-1β, which further confirmed the anti-inflammatory functions of cornuside.
NF-κB signaling is crucial during the development of DN [25]. NF-κB inhibitor alpha (IκB-α) is an important inflammation mediator, which modulates the activation of p50/p65 heterodimer, and then the p50/p65 subunits consequentially translocate into the nucleus and bind with the target inflammatory genes upon IκB-α degradation [26]. In addition, the IκB-α/NF-κB is actively involved in the progression of DN [27]. In this study, we found that cornuside reduced the translocation of activated NF-κB to the nucleus and also inhibited the phosphorylation of IκB-α suggesting that cornuside suppressed the IκB-α/NF-κB signaling pathway during DN progression.
Recent studies have shown that inflammation and ECM accumulation play important roles in the progression of DN. Chronic hyperglycemia may increase the levels of AGEs [28], while AGEs increase the production of transforming growth factor-1 (TGF-β1) in glomerular cells, which can lead to abnormal production of ECMs, inducing glomerular sclerosis and interstitial tubule damage [29]. Therefore, the inhibition of ECM protein production may be an effective method for the prevention and treatment of DN. Our results showed that cornuside inhibited HG-induced overexpression of COL1 and fibronectin. Furthermore, previous studies have shown that an increase in AGEs can stimulate NF-κB signaling, leading to an increase in ECM synthesis [30]. Our results suggest that cornuside may relieve DN by inhibiting NF-κB signaling, which, in turn, decreases ECM synthesis and improves DN symptoms.
PI3K/AKT signaling plays an important role in inflammation-mediated diseases, including DN [31-33]. For instance, microRNA-126 may inhibit inflammation and apoptosis in DN via regulating the PI3K/AKT signaling pathway [34]. Furthermore, tetramethylpyrazine alleviates DN through activating AKT signaling [35]. Herein, we evaluated the effects of cornuside on the expression of key molecules involved in the AKT signaling pathway. We found that cornuside decreased the expression of PCNA, cyclin D1, and p-AKT and increased the expression of p21, implying that cornuside inhibited the AKT signaling pathway during the progression of DN.
In summary, the results of this study indicate that cornuside can inhibit the proliferation of high glucose-treated MMCs and alleviate the inflammatory condition through suppressing the AKT and NF-κB signaling pathways. Therefore, the results of this study demonstrate that cornuside can serve as an alternative medication for the treatment of DN.

 XML Download
XML Download