

 PDF
PDF Citation
Citation Print
Print



Introduction
Toll-like receptor (TLR) family comprises 10 members in humans and 13 in mice [1] whose expression ranges from immune and non-immune cells [2, 3]. TLRs are able to recognize a broad repertoire of ligands defined pathogen-, microbiome- or damage-associated molecular patterns [4]. After recognition of their cognate ligands, TLRs activate the immune response to maintain tissue homeostasis and for host defence [2]. Moreover, each TLR differs in cellular and subcellular localization, reflecting their specificity of their functions [5]. For example, TLR4 is expressed on the cell membrane and recognizes lipopolysaccharide of Gram-negative bacteria [6] while TLR7 is localized on the endosomal compartment and interacts with single-stranded RNAs molecules [7].
Beside the well-known role of TLRs in triggering the immune response, several lines of evidence highlight their involvement in regulating morphogenesis in mammalian and non-mammalian species [8, 9], as testified by several publications in the field [9-13].
We have previously described TLR4 and TLR7 expression pattern during mouse dorsal root ganglia, paravertebral ganglia, and enteric nervous system development, describing a highly-regulated spatiotemporal appearance of these two TLRs [14].
Considering that TLR4 and TLR7 expression is not restricted to the nervous system and the gastrointestinal tube [15, 16], in the present study we extended our investigation by evaluating the localization and appearance of these TLRs in developing respiratory system, liver and pancreas collected at different stages of mouse embryo development.
Materials and Methods
Animals
CD1 mice (Charles River, Calco, Italy) were housed in a thermostatically maintained animal house (T=22°C±2°C; relative humidity 55%±5%) with a 12 hours light cycle, free food access (Italiana Mangimi) and water ad libitum. Mating was arranged by caging females with one male of proven fertility overnight. The morning with evident vaginal plugs was considered day 0 of gestation (E0). Pregnant mice were euthanized by CO2 inhalation and uteri were removed and kept in ice-cold phosphate-buffered saline (PBS) 0.1 M pH 7.4 for dissection and explant of conceptuses at different developmental stages (from E12 to E18). Since respiratory system, liver and pancreas begin to appear in their peculiar morphology at E12 [17], this developmental stage was selected as earliest time point for the subsequent described analyses. The animal protocol was approved by the Ministry of Health – Department for Veterinary Public Health, Nutrition and Food Safety Committee. Animals were treated humanely and in compliance with procedures that alleviate suffering.
Histology and immunohistochemistry staining
Samples were processed as previously reported [14, 18, 19]. Eexcept E12 embryos, processed as a whole, mouse conceptuses were subdivided in head, upper limb and lower limb portions, 4% paraformaldehyde immersion-fixed, dehydrated with ascending grades of ethanol, cleared in xylene, and paraffin embedded. Tissue sections (4 µm thick) were stained with hematoxylin-eosin (H&E) and Masson’s trichrome for histological examination [20].
To evaluate TLR4 and TLR7 expression, immunohistochemistry was performed as previously described [18]. Antigen retrieval was performed on deparaffinized sections by autoclave (6 minutes at 120°C in Na citrate buffer 0.01 M pH 6). Sections were incubated with a solution of 0.3% H2O2 in PBS for endogenous peroxidase activity quenching. Non-specific binding site saturation was performed for 30 minutes with a solution of 0.05 M Tris-HCl, 0.15 M NaCl, 0.1% gelatin, 0.5% ovalbumin, 0.05% Tween-20, and 0.2% fish gelatin (all from Merck). Sections were then incubated with polyclonal rabbit anti-mouse TLR4 (dilution 1:50; overnight at 4°C; Novus Biologicals) or rabbit anti-mouse TLR7 (dilution 1:100; 1 hour at 37°C; Imgenex). Goat anti-rabbit antibody (dilution 1:100; 1 hour at room temperature, Sigma-Aldrich), rabbit peroxidase anti-peroxidase antibody (PAP; DAKO, dilution 1:100; 1 hour at room temperature) were utilized as secondary antibodies. Liquid DAB substrate chromogen system (DAKO; Agilent) were used to develop the reaction. To evaluate the distribution and localization of neurons and nerve fibers, β-tubulin (mouse anti mouse-β3-tubulin, dilution 1:200; 1 hour at 37°C; Santa Cruz Biotechnology) immunohistochemistry was performed. Slides were then dehydrated through a series of alcohol solutions in ascending concentrations, counterstained with hematoxylin, mounted with Entellan (Merck). Samples were observed under a Nikon Eclipse 80i microscope (Nikon) equipped with a digital Nikon DS-L1 camera. The omission of the primary antibody, the sequential omission of the secondary antibody and PAP, or the incubation with DAB alone to exclude the possibility of non-suppressed endogenous peroxidases served as internal controls. Immunohistochemical experiments were performed on at least ten paraffin sections for each sample.
Results
Graphic representation and morphological analysis of mouse embryos at different gestational stages in the respiratory and digestive apparatus
The observation of sections obtained from mouse conceptuses at different developmental stages confirmed the morphological preservation of samples and showed the regions considered for TLR4 and TLR7 immunohistochemical analysis of the respiratory system, liver and pancreas. (Supplementary Figs. 1, 2).
TLR4 expression in the developing respiratory apparatus, liver and pancreas of mouse embryos at different gestational stages
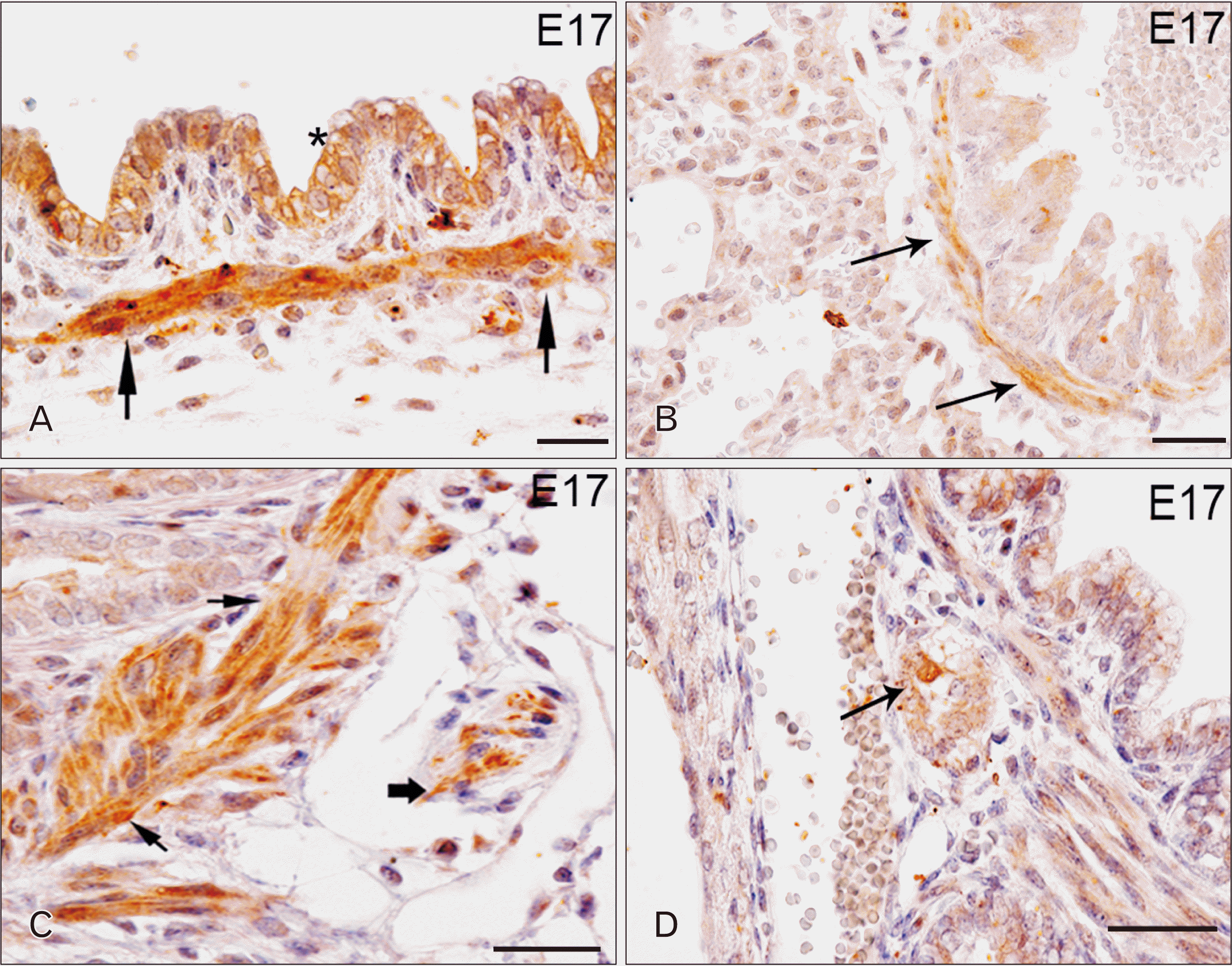
During the development of the respiratory system, TLR4 immunostaining begins to be detectable at E17 in the cytoplasm of cells belonging to the tracheal pseudostratified epithelium (Fig. 1A, asterisk) and in tracheal smooth muscle cells (Fig. 1A, arrows). At the same time-point, TLR4 immunoreactivity was also found in intrapulmonary bronchi (Fig. 1B, C, arrows) and in small ganglia located in the wall of intrapulmonary bronchi (Fig. 1C, thick arrow and Fig. 1D, arrow). Moreover, TLR4 positive cells were observed in bronchioles and in nerve fibers running in the pulmonary parenchyma (data not shown). At E18 stage, TLR4 expression remains unchanged (data not shown).
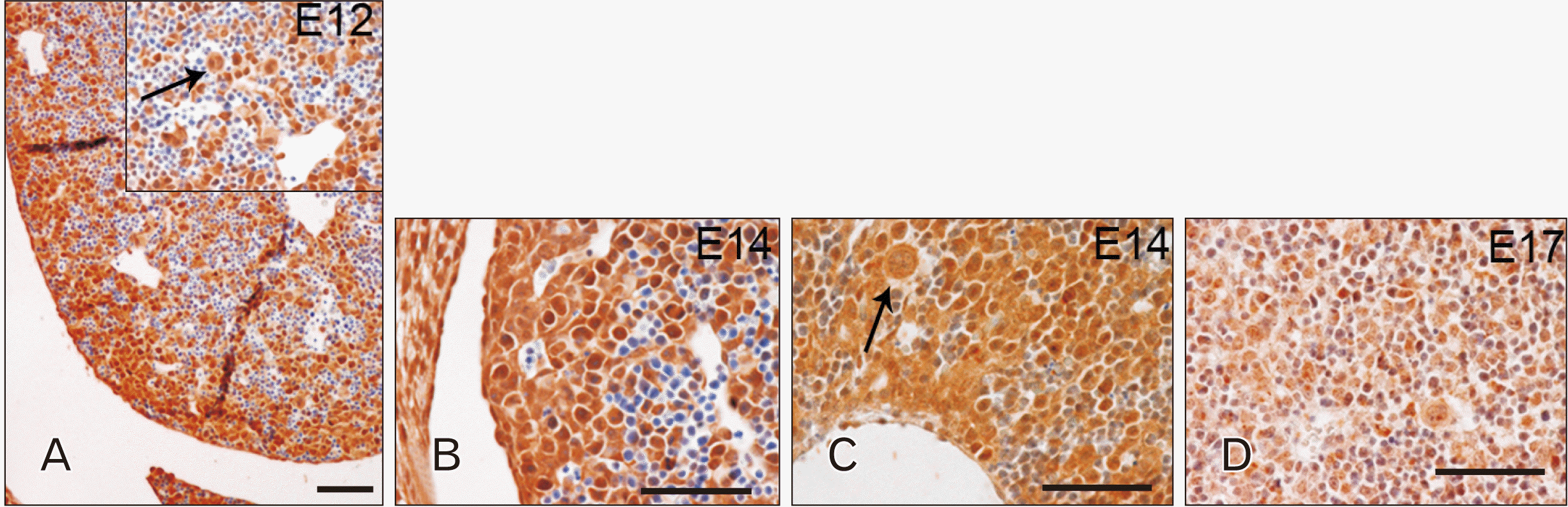
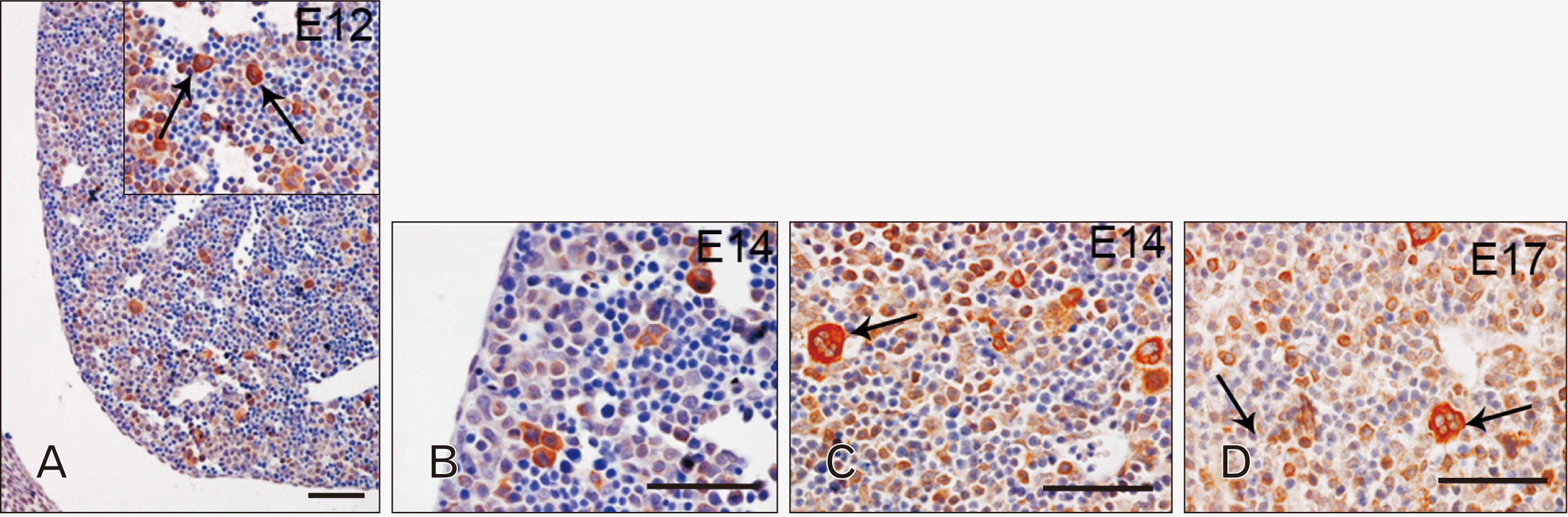
A completely different spatiotemporal pattern of expression was observed when analysing the liver. Starting from E12, TLR4 appeared to be localized in cells of hepatoblast derivation, particularly abundant at the periphery of the organ (Fig. 2A). Cells differentiated from erythroblast progenitors were found TLR4 negative, while the cytoplasm of differentiating megakaryocytes was found TLR4 positive (Fig. 2A, arrow in the inset). TLR4 distribution pattern was maintained at E14 (Fig. 2B, C), while at E17 its expression is weaker (Fig. 2D).
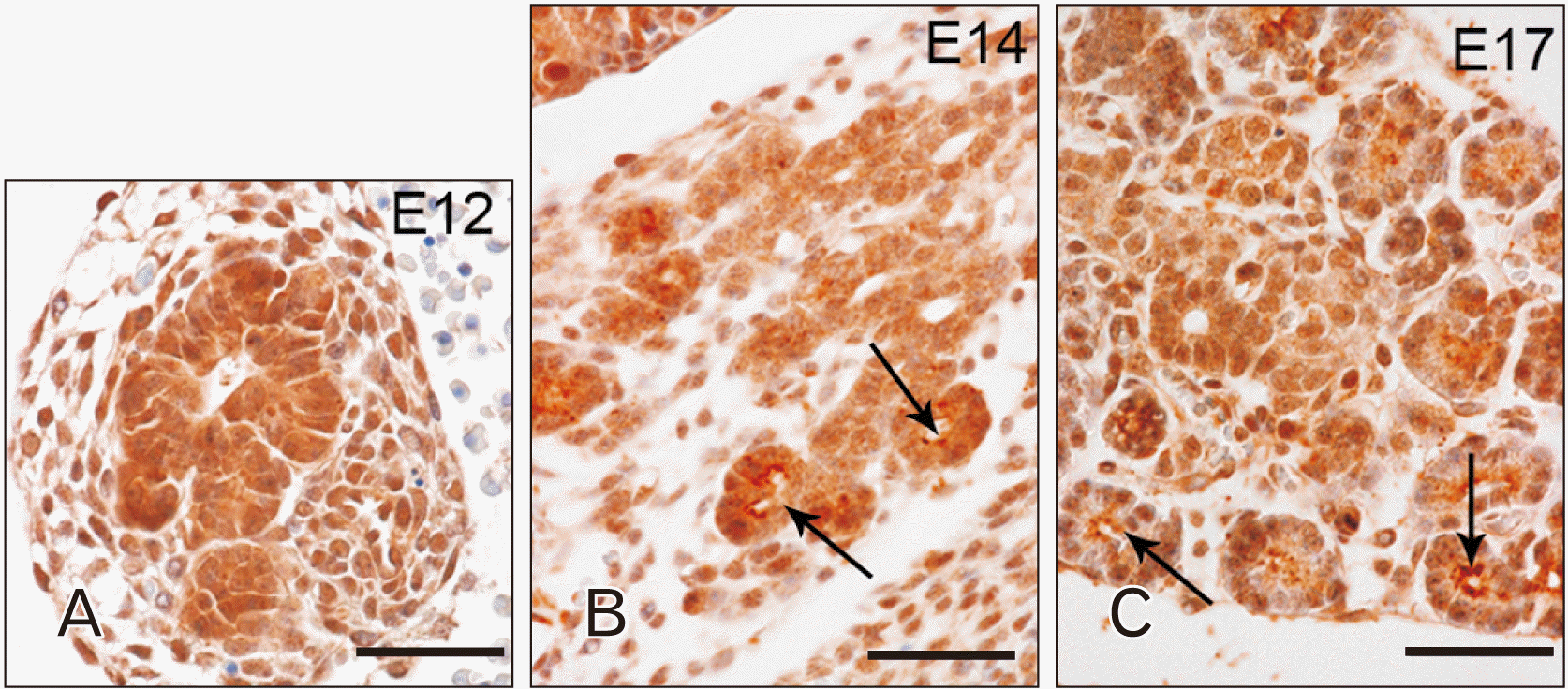
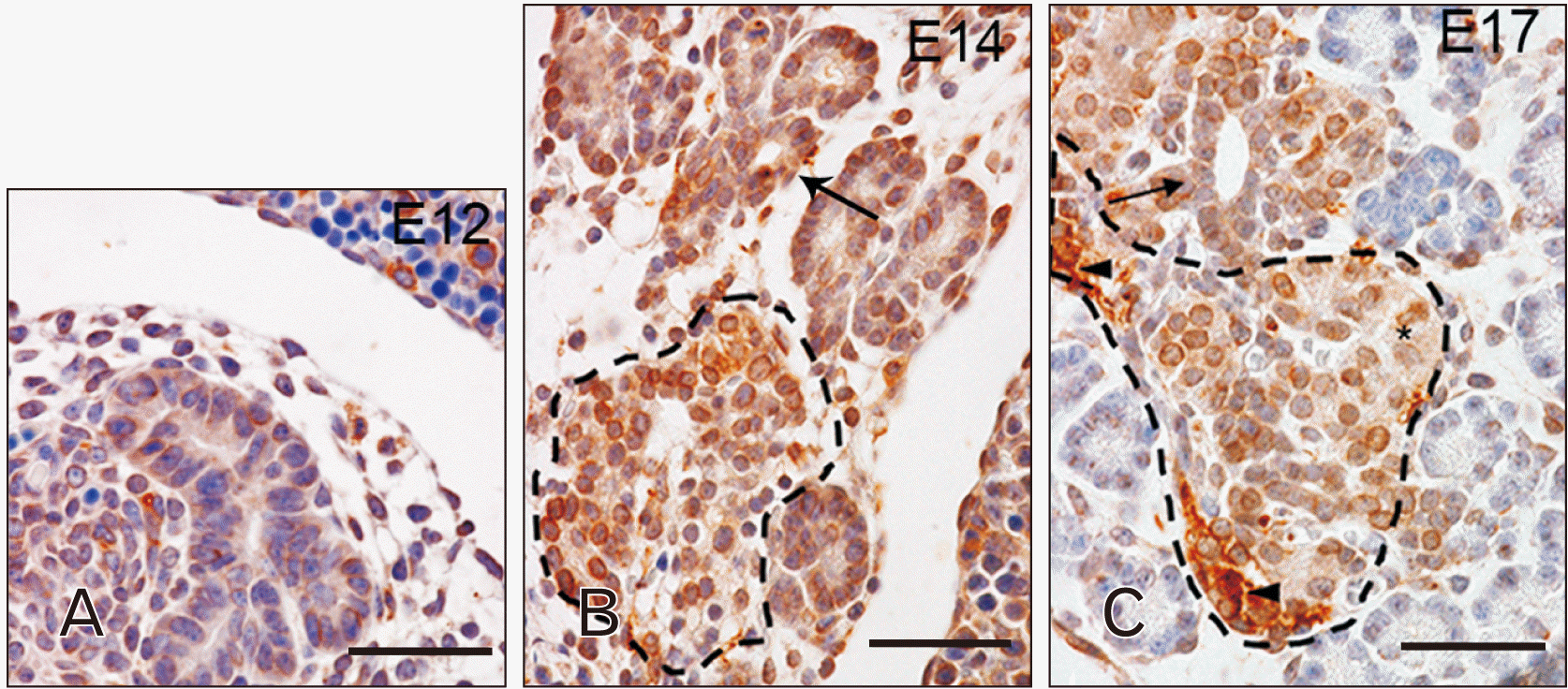
From E12, TLR4 appeared in cells of the ventral and dorsal pancreatic primordial buds (Fig. 3A). At this embryonic stage, TLR4 showed cytoplasmatic localization in almost all the cells of primordial buds while, at E14, it can be possible to observe this TLR in the apical area of cells in the developing exocrine acini (Fig. 3B, arrows). At E17, TLR4 localization remained unaltered (Fig. 3C, arrows).
TLR7 expression in the developing respiratory apparatus, liver and pancreas of mouse embryos at different gestational stages
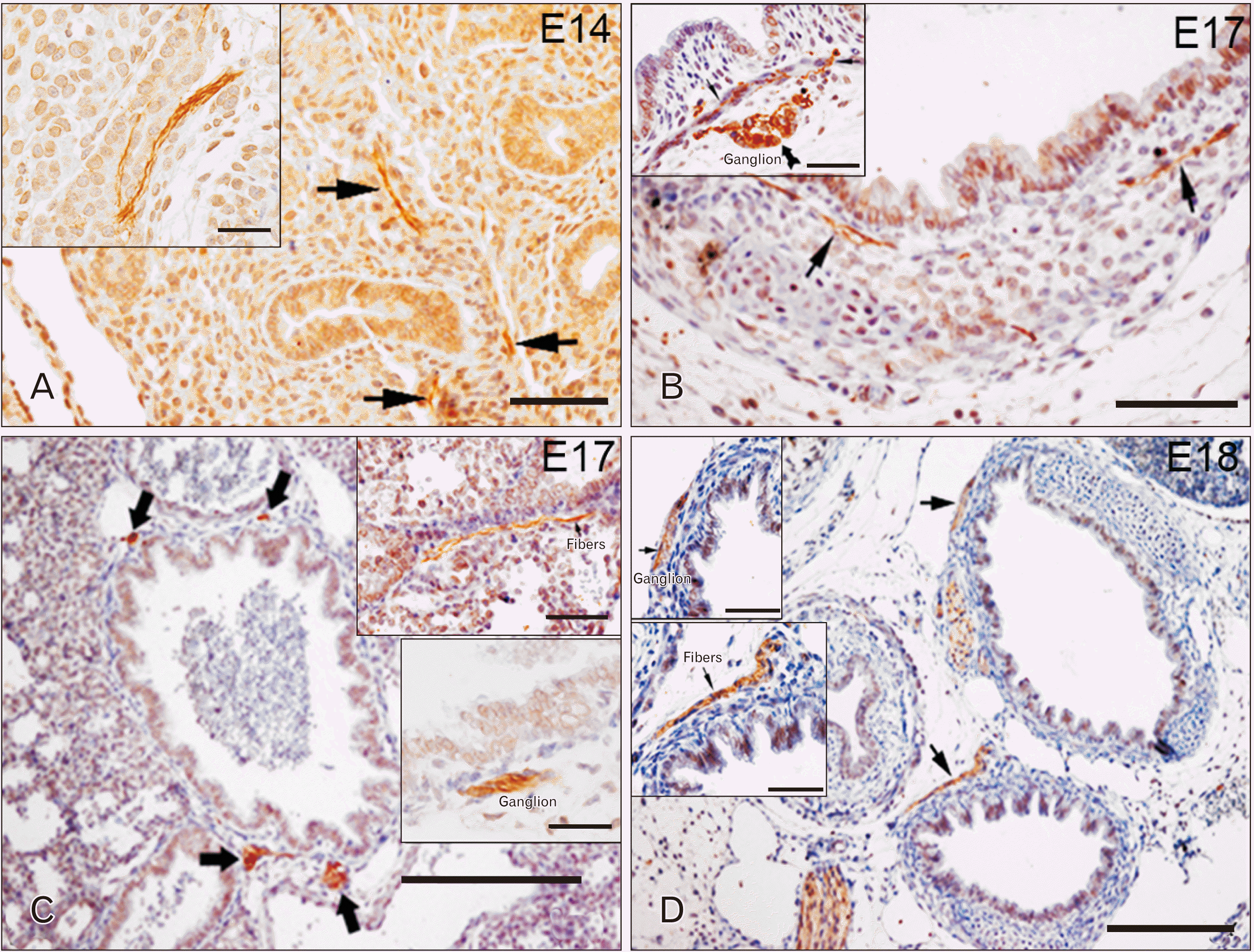
TLR7 was detectable in the developing respiratory system starting from E14, especially localized in some nerve fibers running in the lung parenchyma (Fig. 4A, arrows). At E17, TLR7 appeared also in small ganglia and nerve fibers in the smooth muscle cells of the tracheal wall (Fig. 4B, thick arrow in the inset and arrows, respectively), pulmonary parenchyma and bronchioles (Fig. 4C, thick arrows). At E18, in addition to the same structures mentioned before, TLR7 immunoreactivity was also observed in small ganglia and in nervous fibers of the smooth muscle of main bronchi (Fig. 4D).
In the developing liver, TLR7 expression pattern was similar to that observed for TLR4 at every considered embryological stage, but always exhibiting a weaker intensity. At E12, TLR7 positivity was found stronger in megakaryocytes than in cells of hepatoblast derivation (Fig. 5A, arrows in the inset indicate megakaryocytes). Starting from E14, TLR7 expression appeared also in developing hepatocytes and cholangiocytes, especially localized close to the centrilobular veins (Fig. 5B, C). Similar to TLR4, TLR7 staining became weaker by E17 (Fig. 5D).
In developing pancreas, TLR4 and TLR7 expression was detected in cells of the ventral and dorsal pancreatic primordial buds at E12 (Fig. 6A). At this embryonic stage, TLR7 was evident in the perinuclear region of a few number of cells (Fig. 6A). At E14, TLR7 labelling extended to nearly all cells of the developing pancreas, appearing more evident in the region of ducts and endocrine parenchyma, such as the islets of Langerhans than in the region of exocrine acini (Fig. 6B, arrow and dotted line, respectively). At E17, TLR7 immunoreactivity remained clearly visible only in the cells forming the excretory ducts and islets of Langerhans, showing a perinuclear and cytoplasmic localization (Fig. 6C, arrows and dotted line, respectively).
Discussion
In the present work, we evaluated the spatiotemporal pattern of expression of TLR4 and TLR7 during mouse respiratory system, liver and pancreas development, all organs that, directly or indirectly, are in communication with the external environment. Although the respiratory and digestive apparatus arise from endodermal derivatives [21], we observed that TLR expression during embryo development is not homogeneous either among different systems and among the different tissues constituting single organs, possibly suggesting the participation of these TLRs in the formation of the different body structures.
The pattern of staining in the liver parenchyma, especially for TLR4, seems to mirror the maturation of the organ itself. Indeed, the majority of TLR4 positive cells, of possible hepatoblast derivation, was initially localized at the periphery of the liver whereas, at a later time point (E14), it was possible to appreciate an increased presence of immunoreactive elements around the central veins. Finally, a decline in staining intensity was observable at E16. Therefore, the expression of TLR4 in the hepatoblast component may embrace the period of initial structuring of the liver parenchyma and decline during the liver transition from fetal hematopoietic organ to adult exocrine/endocrine gland. A possible explanation of this expression pattern might rely on the role played by TLRs in proliferation, differentiation and survival of hematopoietic progenitor cells [22, 23]. For instance, it has been reported that TLR signalling pathway can orchestrate hematopoietic progenitor cells differentiation towards the myeloid lineage, suggesting a possible involvement of these receptors in the development of blood/immune cells occurring in the foetal liver [23, 24].
In the embryo pancreas, at E12, TLR4 intensity staining was localized in all the pancreatic primordium whereas TLR7 was only present in scattered cells. At later stages of development, the time of appearance of these two receptors seems to be strictly related to the time of differentiation of primordial cells toward an exocrine or endocrine fate [25]. Indeed, TLR4 is mainly expressed in the exocrine component of the organ, especially in differentiating acini, while TLR7 was found in the area within the ducts where the developing endocrine cells are located. It is possible to speculate that the ability of these TLRs to bind elements present in the extracellular environment might be involved in the pancreatic primordial cell fate determination. Indeed, TLRs are reported to interact with extracellular matrix (ECM) proteins or molecules involved in cell adhesion and to modulate metalloproteinase activity [26-30] and, accordingly, matrix degradation and remodelling is crucial for epithelial morphogenesis and for the endocrine precursors differentiation and migration [31]. Therefore, it is possible to depict a scenario in which TLRs are deputed at recognizing ligands released during embryonic tissue rearrangements or in mediating cell-cell contact and, subsequently, triggering a signalling cascade that determines a molecular reprogramming of the cells that are expressing those receptors.
In the airways, the temporal sequence of expression for TLR4 (E17) and TLR7 (E14) and the localization of these two receptors appears to be similar to that previously observed during gut development [14], probably because these two systems are connected with the external environment that allows a rapid bacteria colonization immediately after birth. As in the intestine, the expression of TLR4 in the respiratory system is detectable only from E17 in the smooth muscle and in the intrinsic nervous structures, suggesting that the prenatal expression of the TLR4 receptor in the airways may be not so strictly connected to a morphogenetic role but, instead, with its well-known involvement of TLRs in the immune responses. Another intriguing possibility is that the late expression of TLR4 during embryo development may be linked to the initiation of parturition. Indeed, it has been demonstrated that surfactant protein secreted by mouse foetal lungs can trigger the delivery process [32]. Since TLR4 is able to recognize surfactant protein A [33], it is possible the a late-stage appearance of this TLR may prevent unwanted preterm labour. Moreover, it cannot be excluded that TLR4 expression may be substantially linked to the processes of maturation of the immune responses in preparation for the passage from the intra-uterine to the extra-uterine environment.
In the case of the TLR7, its expression in the nervous component of the airways is clearly visible starting from the E14 stage in ganglia and nerve fibers running in the lung parenchyma. The expression of TLRs in the developing peripheral nervous system, particularly at early stages, is not restricted to their immunological function. Indeed, it should be remembered that in Drosophila, the protein Toll possesses a fundamental morphogenetic role of the nervous system including synaptogenesis and axonogenesis [34-38]. Although still to be fully elucidated, it appears that TLRs, mammalian orthologs of the above-mentioned protein, can maintain an active involvement in determining the fate of cells derived from the neural crest, from which a large part of the peripheral nervous system originates [39], as demonstrated for some TLR family members [9, 40, 41].
Another possible view is based on the fact that, at least starting from the foetal period, microbes start colonizing gut mucosal surfaces creating bacterial communities defined as microbiota [42]. Microbiota not only plays a fundamental role in maintaining a sort of immunological homeostasis of the tissues in which it is residing but also concurs to ensure a good health status of the entire body, as testified by several studies in this field [43, 44]. Although the majority of microorganisms resides in the intestine, it is emerging that other organs, historically considered “sterile” such as lungs and pancreas, are characterized by the presence of resident bacteria [45-47]. For instance, in lungs commensal bacteria contribute to create a mildly immunosuppressed environment able to prevent inflammatory responses caused by the inhalation of particulates present in the air [45].
Since it is now well known that TLRs are important characters in the “dialog” between host cells and the microbiota [48], the appearance of these receptors during development may be crucial to guarantee an adequate and immediate recognition of these commensals as soon as they colonize the body.
 XML Download
XML Download