

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Myocardial ischemia and reperfusion injury (MIRI) is the pathological process leading to the death of cardiomyocytes, which significantly changes heart function after infarction and increases heart failure risk.1) MIRI induces sterile injury and inflammation and is characterized by the release of DAMPs, thereby initiating and triggering NOD-like receptor pyrin domain-containing protein 3 (NLRP3) inflammasomes.2) Despite advances in cardiovascular medicine, MIRI still increases mortality and lengthens hospital stays. Therefore, understanding the MIRI molecular mechanism will help to find a treatment strategy that ultimately reduces the infarct size.
Pyroptosis is an innate inflammatory response caused by different pathological irritation, including stroke, heart disease, or cancer, and is essential for commanding microbial infections. It is characterized by serosal membrane rupture, which releases intracellular and proinflammatory contents.3) It mainly causes gasdermin D (GSDMD) division and cytokine activation, including interleukin (IL)-18 and IL-1β.4) The NLRP3 inflammasome activation-mediated pyroptosis pathway is related to MIRI. A previous study reported that metformin protected MIRI and pyroptosis through the AMPK/NLRP3 inflammasome pathway.5) Ye et al.6) reported that emodin reduced MIRI by inhibiting GSDMD-mediated cardiomyocyte pyroptosis. The above showed that pyroptosis played a vital role in MIRI. However, the pyroptosis molecular mechanism in MIRI needs to be further explored.
In the past decade, long noncoding RNAs (lncRNAs) have attracted much attention. The size of lncRNAs exceeds 200 nt, and a series of significant life progression, including cell proliferation, differentiation, and apoptosis, can be regulated by lncRNAs.7) Lysyl oxidase like 1 antisense RNA 1 (LOXL1-AS1) is an lncRNA that plays a vital role in several diseases. For instance, LOXL1-AS1 facilitated gastric cancer occurrence and stemness via regulating the microRNA (miRNA)-708-5p/USF1 pathway.8) Notably, Song et al.9) reported that the LOXL1-AS1 level was upregulated in patients with coronary heart disease. LOXL1-AS1 enhanced oxidized low-density lipoprotein and caused human coronary artery endothelial cell (HCAEC) pyroptosis. The abovementioned studies suggested that LOXL1-AS1 played a regulatory role in cardiovascular diseases by modulating cell pyroptosis. However, the functional role of LOXL1-AS1 in MIRI is unclear.
MiRNAs are a kind of endogenous small RNAs, which regulate gene expression after transcription. MiRNA regulates its target gene through translation inhibition and mRNA destabilization.10) A previous study showed that the knockdown of miR-29a inhibited oxidative stress and NLRP3-mediated pyroptosis by targeting SIRT1 in MIRI.11) Zhou et al.12) reported that miR-132 promoted oxidative stress-induced pyroptosis in MIRI by targeting sirtuin 1. MiR-761 is closely associated with cardiovascular disease progression. A previous study reported that miR-761 modulated macrophage-derived foam cell formation and inflammation in atherosclerosis.13) Furthermore, miR-761 could inhibit apoptosis and protect mouse cardiomyocytes from H/R-induced damage.14) Notably, miR-761 could repress pyroptosis-associated protein IL-1β and IL-18 levels.15) Additionally, through bioinformatics, we predicted that LOXL1-AS1 and miR-761, miR-761 and phosphatase and tensin homolog (PTEN) had mutual binding sites, but their regulatory mechanisms in MIRI were still unclear.
Based on the above background, we made a reasonable hypothesis that LOXL1-AS1 may regulate PTEN expression by targeting miR-761 to enhance cardiomyocyte pyroptosis and promote MIRI. Our study may have predictive and preventive values for MIRI.
METHODS
Ethical statement
Animal studies were approved by the Ethics Committee of the First People’s Hospital of Chenzhou (No. 2020031).
Cell culture and treatment
The human cardiomyocyte (HCM) cell line was purchased from the Shanghai Xuanke Biotechnology Co., Ltd (Shanghai, China). HCMs were cultured in Dulbecco’s Modified Eagle’s Medium (DMEM) with 10% fetal bovine serum. To simulate in vitro myocardial H/R injury, HCMs were treated with reperfusion after hypoxia and glucose deprivation. HCMs were exposed to glucose-free DMEM and cultured at 95% N2 and 5% CO2 conditions. After culturing for 4 hours, the cells were put back into the normal glucose-containing medium (4.5 mg/mL) and reoxidized for 24 hours under normal 95% air and 5% CO2 conditions for subsequent experimental testing.
Plasmid construction and transfection
Short hairpin RNA of LOXL1-AS1 (sh-LOXL1-AS1, 5′-GGTGCATACAAGCACTCAATA-3′), miR-761 mimics (5′-GCAGCAGGGUGAAACUGACACA-3′), miR-761 inhibitor (5′-UGUGUCAGUUUCACCCUGCUGC-3′), and their negative control vectors (sh-NC: GTTCTCCGAACGTGTCACGT, mimics NC: 5′-UUGUACUACACAAAAGUACU-3′, inhibitor NC: 5′-CAGUACUUUUGUGUAGUACAA-3′) were synthesized and packed in lentiviral vectors by GenePharma (Shanghai, China). When grown to 50–60% confluence, cells were washed with phosphate buffer solution and changed to a serum-free medium. Subsequently, the cells were transfected with these lentivirus vectors in the presence of 10 μg/mL polybrene and selected using puromycin for 1 week. Lipofectamine 2000 was utilized to perform cell transfection following the manufacturer’s instructions. Subsequently, RNA was collected for quantitative real-time polymerase chain reaction (qRT-PCR) to verify the knockdown efficiency after transfection for 48 hours.
Animal model
Thirty-five adult male C57BL/6 mice were randomly divided into the control, sham, ischemia/reperfusion (I/R), I/R+sh-NC, and I/R+sh-LOXL1-AS1 groups, with 5 mice in each group. The mice were anesthetized by intraperitoneal injection of pentobarbital sodium before tracheal intubation. The ventilator parameters of anesthetized mice were synchronously set with the breathing rate. Subsequently, the endotracheal tube was connected to the ventilator. The chest of the mouse was opened from the third intercostal space at the left margin of the sternum. Next, the left anterior descending coronary artery was ligated using a 7-0 silk suture. The ligature was loosened for reperfusion 45 minutes later, the heart tissue was collected 24 hours later, and blood was taken from the heart before sacrifice. Mice in the control group did not undergo any treatment. The mice in the sham group had their chest open, and the corresponding parts were threaded without ligation. A volume of 10-μL concentrated lentivirus with sh-LOXL1-AS1 or sh-NC (1×109 PFU/mL) was delivered via intramyocardial injection into the apex and anterolateral wall 48 hours before modeling.
Biochemical detection
The corresponding commercial kits were used to test the levels of lactate dehydrogenase (LDH), creatine kinase-MB (CK-MB), and cardiac troponin I (cTnI) according to the manufacturer’s instructions (Nanjing Jiancheng Bioengineering Institute, Nanjing, China). Multiskan SkyHigh microplate reader (Thermo Fisher Scientific, Waltham, MA, USA) was used to calculate the LDH, CK-MB, and cTnI levels by forming the standard curve with provided values.
Enzyme-linked immunosorbent assay
IL-18 and IL-1β levels in the serum of mice and cell supernatant were evaluated using enzyme-linked immunosorbent assay (ELISA). IL-18 (CUSABIO, Wuhan, China) and IL-1β (CUSABIO) were quantitated according to the manufacturer’s instructions. Multiskan SkyHigh microplate reader (Thermo Fisher Scientific) was used to calculate IL-18 and IL-1β concentrations by forming the standard curve with provided values.
MTT
MTT was used to characterize cell viability. Cells at the logarithmic growth stage were digested and counted. The cells were inoculated into 96-well plates at a density of 1×104 cells/well. Subsequently, the cells were added with 5 mg/mL MTT (10 μL; Sigma-Aldrich, St. Louis, MO, USA) and incubated for 4 hours at 37°C with 5% CO2. Next, 96-well plates were taken out, the medium containing MTT was sucked out, and 150-μL DMSO was added to each well. After shaking for 10 minutes, the absorbance value (490 nm) was analyzed using a microplate reader.
Quantitative real-time polymerase chain reaction
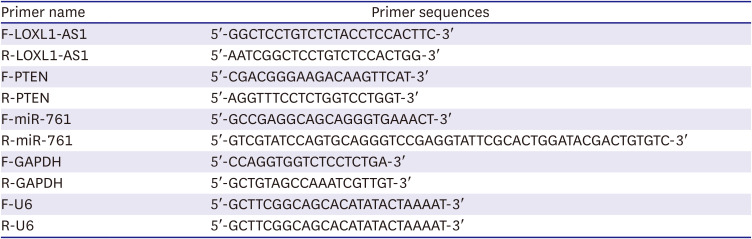
LOXL1-AS1, miR-761, and PTEN levels in cells and tissues were measured using qRT-PCR. Briefly, total RNA was extracted by Trizol reagent (Invitrogen, Carlsbad, CA, USA). RNA quality and quantity were assessed with agarose gel electrophoresis and A260/A280 ratio with spectrophotometer. The A260/A280 ratios for all RNA preparations were about 1.8–2.0. For miRNA, a miRNA reverse transcription kit (CW2141; CWBIO, Beijing. China) was used to synthesize cDNA. For mRNA, an mRNA reverse transcription kit (CW2569; CWBIO) was applied. Ultra SYBR Mixture (Thermo Fisher Scientific) was applied to test the gene level, which was calculated using the 2−ΔΔCt method. The reaction parameters set by the PCR instrument as follows: 95°C for 10 minutes followed by 45 cycles consisting of 95°C for 15 seconds, 58°C for 30 seconds and 68°C for 60 seconds. U6 and glyceraldehyde-3-phosphate dehydrogenase (GAPDH) were used as the internal reference for miRNA and mRNA, respectively. Primer sequences are shown in Table 1.
Table 1
Primer sequences
Western blot
To extract the total protein in cells and tissues, RIPA lysis buffer was employed. Protein was quantified in each group using the BCA kit (Biosharp, Anhui, China) and subsequently mixed with sodium dodecyl sulfate polyacrylamide gel electrophoresis buffer; the protein was adsorbed on the polyvinylidene fluoride membrane. Then, the membranes were blocked in 5% bovine serum albumin (BSA) for 1 hour. ASC (10500-1-AP; Proteintech, Rosemont, IL, USA), cleaved caspase-1 (24232; CST, Danvers, MA, USA), GSDMD-N (ab215203; Abcam, Cambridge, UK), NLRP3 (19771-1-AP; Proteintech), IL-18 (10663-1-AP; Proteintech), IL-1β (16806-1-AP; Proteintech), PTEN (9188; CST), and GAPDH (AB9485; Abcam) primary antibodies were used for overnight incubation. We then incubated horseradish peroxidase (HRP)-labeled secondary antibody IgG-HRP (BL003A; BioSharp). Exposure was performed using an ECL chemiluminescence substrate (Biosharp). To measure protein expressions, GAPDH was used as an internal reference.
Flow cytometry
Cell pyroptosis was measured by flow cytometry. The cells were digested collected, and centrifuged at 2,000 rpm for 5 minutes, and approximately 3×105 cells were collected. FAM-YVAD-FMK (FAM600-1; Cell Technology, Hayward, CA, USA), caspase-1 antibody, and propidium iodide were added and mixed. Subsequently, these were incubated at normal temperature and in the dark for 1 hours. Flow cytometry (Beckman, CytoFLEX, Bria, CA, USA) was subsequently performed. Data were measured using a FACSCanto II flow cytometer system, and flow cytometry results were analyzed using Flowjo software (TreeStar, San Francisco, CA, USA).
Immunofluorescence
HCMs were fixed with 4% paraformaldehyde. Then, 0.3% tretinoin was added and permeabilized. HCMs were blocked at 37°C with 5% BSA for 1 hour. The primary antibody, NLRP3, was added overnight. Subsequently, a secondary antibody was added and incubated at 37°C. Nuclei were stained with DAPI. Slices were blocked in buffered glycerol and observed using fluorescence microscopy.
Dual-luciferase reporter gene assay
The Starbase database (http://starbase.sysu.edu.cn) was used to predict the binding relationship among LOXL1-AS1, miR-761, and PTEN. Wild-type (WT) or mutant type (MUT) of LOXL1-AS1 and PTEN fragments were inserted into the pmirGLO vector using a dual-luciferase assay kit (E1910; Promega, Madison, WI, USA). Recombinant vectors, NC mimics or miR-761 mimics, were transfected into cells using Lipofectamine 2,000 (Thermo Fisher Scientific). Finally, the luciferase activity was measured using a GloMax 20/20 chemiluminescence detector (Promega).
Hematoxylin and eosin staining
Morphological changes in the myocardium were detected using hematoxylin and eosin (HE). The mice cardiac muscle tissues were fixed in 4% paraformaldehyde for 24 hours. After dehydration and paraffin embedding, the tissues were sliced into 5-μm sections using a slicer. The slices were then baked, dewaxed, and stained using HE and dehydrated using gradient alcohol. After taking them out, they were placed in xylene and sealed with neutral gum. The myocardial injury degree was observed using a microscope (BA210T; MOTIC, Xiamen, China).
Immunohistochemistry
The positive rates of NLRP3 and caspase-1 in the myocardium tissues were detected by immunohistochemistry (IHC). The slices were baked and dewaxed, and the antigen was repaired using heat. We then added them into 1% periodate acid for 10 minutes. NLRP3 or caspase-1 primary antibody was incubated overnight at 4°C. The secondary antibody was incubated at 37°C for 30 minutes. DAB, hematoxylin, and alcohol (60–100%) were applied for 5 minutes in sequence. After removal, tablets were soaked in xylene, and the slices were subsequently sealed using neutral adhesive and observed under a microscope.
Statistical analysis
Statistical analysis was performed using GraphPad Prism 8.0 software (GraphPad Software Inc., San Diego, CA, USA). Data were presented as mean±standard deviation. Unpaired t-test was utilized between the 2 groups that fit the normal distribution. One-way analysis of variance and Tukey’s post-hoc test were performed to compare multiple groups. The p<0.05 indicated a statistically significant difference.
RESULTS
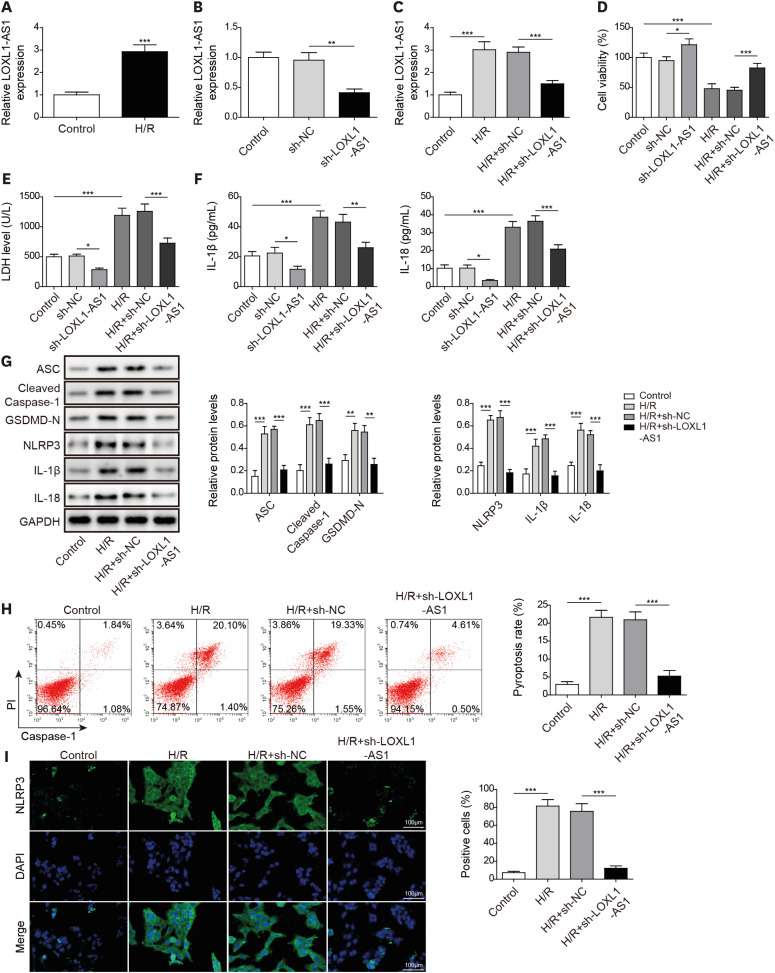
Knocking down long noncoding RNA lysyl oxidase like 1 antisense RNA 1 improved hypoxia/reoxygenation-induced human cardiomyocyte pyroptosis
To examine the LOXL1-AS1 function in H/R-treated cells, we first detected the LOXL1-AS1 level in H/R-induced HCMs. It was observed that LOXL1-AS1 was increased following H/R treatment (Figure 1A). We subsequently used sh-LOXL1-AS1 to knock down LOXL1-AS1 in HCMs (Figure 1B). Furthermore, LOXL1-AS1 levels were facilitated following H/R treatment; sh-LOXL1-AS1 significantly downregulated the LOXL1-AS1 level (Figure 1C). Knockdown of LOXL1-AS1 under basal conditions increased cell viability. H/R treatment inhibited HCM viability, and the knockdown of LOXL1-AS1 could improve the repression in cell viability caused by H/R treatment (Figure 1D). Moreover, sh-LOXL1-AS1 decreased LDH level in both basal conditions and H/R treated conditions (Figure 1E). ELISA result revealed that sh-LOXL1-AS1 treatment inhibited IL-1β and IL-18 release compared with control group, and LOXL1-AS1 knockdown suppressed the release of inflammatory cytokines induced by H/R (Figure 1F). Western blot (WB) further detected the protein level of pyroptosis-related proteins. It was noted that the high levels of ASC, cleaved caspase-1, GSDMD-N, NLRP3, IL-1β, and IL-18 were offset by LOXL1-AS1 knockdown in H/R-treated cells (Figure 1G). Consistently, flow cytometry revealed that H/R promoted cell pyroptosis, and LOXL1-AS1 silence inhibited the severity of H/R treatment-induced pyroptosis (Figure 1H). Additionally, H/R treatment significantly upregulated the number of NLRP3-positive cells, and the knockdown of LOXL1-AS decreased the number of NLRP3-positive cells (Figure 1I). These results demonstrated that LOXL1-AS1 promoted pyroptosis in H/R-induced HCM.
Figure 1
Knocking down LOXL1-AS1 improved H/R-induced human cardiomyocyte pyroptosis. (A) Using qRT-PCR to detect the LOXL1-AS1 level. (B) Using qRT-PCR to examine the LOXL1-AS1 knockdown effect. (C) Using qRT-PCR to test the LOXL1-AS1 level. (D) Using MTT to determine cell viability. (E) Using the Release Assay Kit to monitor the LDH level. (F) Using ELISA to detect IL-1β and IL-18 secretion. (G) Using Western blot to measure ASC, cleaved caspase-1, GSDMD-N, NLRP3, IL-1β, and IL-18 levels. (H) Using flow cytometry to monitor the pyroptosis level. (I) Using immunofluorescence to detect NLRP3 expression. Data were presented as mean±standard deviation, and 3 separate experiments were performed in triplicate.
ELISA = enzyme-linked immunosorbent assay; GAPDH = glyceraldehyde-3-phosphate dehydrogenase; GSDMD = gasdermin D; H/R = hypoxia/reoxygenation; IL = interleukin; LDH = lactate dehydrogenase; LOXL1-AS1 = lysyl oxidase like 1 antisense RNA 1; NLRP3 = NOD-like receptor pyrin domain-containing protein 3; qRT-PCR = quantitative real-time polymerase chain reaction.
*p<0.05; **p<0.01; ***p<0.001.
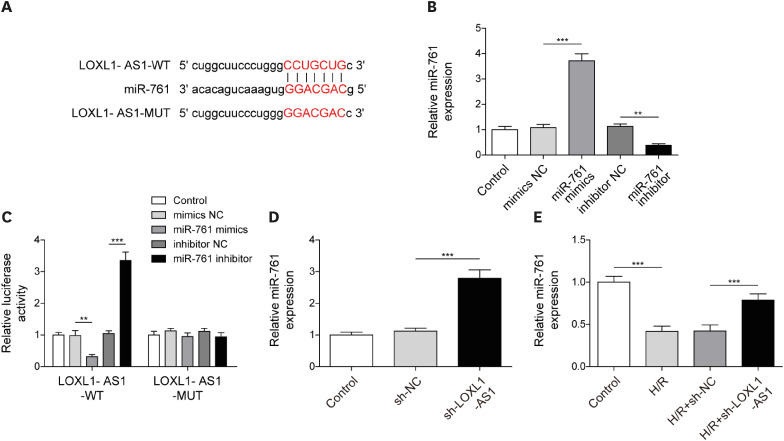
Long noncoding RNA lysyl oxidase like 1 antisense RNA 1 negatively regulated the miR-761 level as a molecular sponge
Subsequently, we predicted the target genes of LOXL1-AS1 using the Starbase database (Supplementary Table 1). Among all the predicted miRNAs, we selected 6 miRNAs of interest and noted that among them, miR-761 was significantly reduced under H/R conditions, and its change was the most significant (Supplementary Figure 1). Through bioinformatics prediction, we observed that LOXL1-AS1 and miR-761 have mutual binding sites (Figure 2A). Then, miR-761 was knocked down or overexpressed by miR-761 inhibitor and mimics, respectively (Figure 2B). Dual-luciferase detection revealed that the miR-761 mimics significantly reduced the luciferase activity of HCMs, and the miR-761 inhibitor significantly enhanced the luciferase activity when cells transfected with WT recombinant LOXL1-AS1 plasmid; however, these treatments had no significant effect on the luciferase activity in the LOXL1-AS1 MUT group, indicating that LOXL1-AS1 had a mutually binding relationship with miR-761 (Figure 2C). Furthermore, the knockdown of LOXL1-AS1 in HCMs increased the miR-761 level (Figure 2D). Moreover, we observed that the knockdown of LOXL1-AS1 had no effect on pri-miR-761 and pre-miR-761 levels (Supplementary Figure 2). Additionally, the miR-761 level was downregulated under H/R conditions, whereas LOXL1-AS1 inhibition upregulated the miR-761 level (Figure 2E). These results indicated that LOXL1-AS1 suppressed miR-761 expression by directly targeting miR-761 in HCMs.
Figure 2
LOXL1-AS1 negatively regulated the miR-761 level as a molecular sponge. (A) Using bioinformatics to predict LOXL1-AS1 and miR-761 binding sites. (B) Using qRT-PCR to examine miR-761 knockdown and overexpression effects. (C) Using dual-luciferase to test LOXL1-AS1 and miR-761 interaction. (D) After interfering with LOXL1-AS1 in HCMs, qRT-PCR is used to monitor the miR-761 level. (E) Under H/R conditions, LOXL1-AS1 was knocked down in HCMs, and qRT-PCR was used to examine the miR-761 level. Data were presented as mean±standard deviation, and 3 separate experiments were performed in triplicate.
H/R = hypoxia/reoxygenation; HCM = human cardiomyocyte; LOXL1-AS1 = lysyl oxidase like 1 antisense RNA 1; MUT = mutant type; qRT-PCR = quantitative real-time polymerase chain reaction; WT = wild-type.
**p<0.01; ***p<0.001.
MiR-761 overexpression improved hypoxia/reoxygenation-induced human cardiomyocyte pyroptosis
We subsequently probed miR-761 function on H/R-induced HCM pyroptosis. MiR-761 mimics markedly increased the miR-761 level in H/R-treated cells (Figure 3A), indicating that miR-761 was successfully overexpressed under H/R. Moreover, the decrease of cell viability and upregulation of LDH level induced by H/R treatment were reversed by miR-761 overexpression (Figure 3B and C). H/R promoted the release of IL-1β and IL-18 and dramatically upregulated the protein level of pyroptosis-related factors; however, those upregulating effects were recovered by miR-761 overexpression (Figure 3D and E). Flow cytometry further confirmed that miR-761 overexpression inhibited the pyroptosis level of H/R-treated cells (Figure 3F). Additionally, H/R treatment significantly upregulated the fluorescence intensity of NLRP3, and miR-761 overexpression improved the upregulation effect of H/R treatment (Figure 3G). These results showed that miR-761 inhibited pyroptosis in H/R-induced HCMs.
Figure 3
MiR-761 overexpression improved H/R-induced human cardiomyocyte pyroptosis. (A) Using quantitative real-time polymerase chain reaction to examine the miR-761 level. (B) Using MTT to examine cell viability. (C) Using the Release Assay Kit to detect the LDH level. (D) Using enzyme-linked immunosorbent assay to determine IL-1β and IL-18 levels. (E) Using Western blot to monitor ASC, cleaved caspase-1, GSDMD-N, NLRP3, IL-1β, and IL-18 levels. (F) Using flow cytometry to detect caspase-1 activity. (G) Using immunofluorescence to detect NLRP3 expression. Data were presented as mean±standard deviation, and 3 separate experiments were performed in triplicate.
GAPDH = glyceraldehyde-3-phosphate dehydrogenase; GSDMD = gasdermin D; HR = hypoxia/reoxygenation; IL = interleukin; LDH = lactate dehydrogenase; NLRP3 = NOD-like receptor pyrin domain-containing protein 3.
*p<0.05; **p<0.01; ***p<0.001.
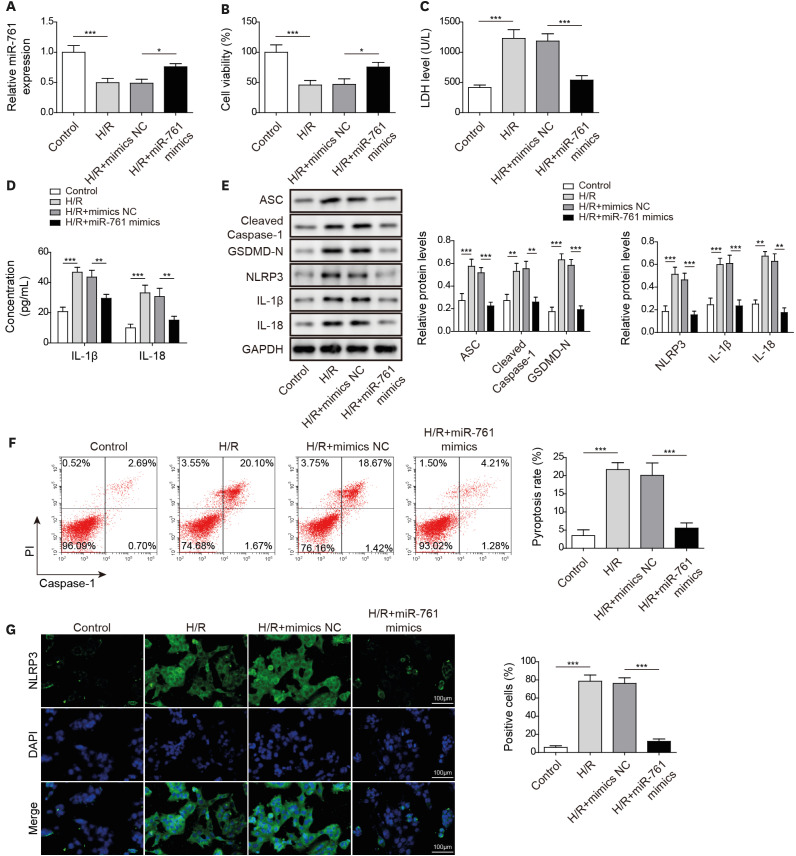
Long noncoding RNA lysyl oxidase like 1 antisense RNA 1 induced human cardiomyocyte pyroptosis by targeting miR-761 under hypoxia/reoxygenation conditions
To further explore the LOXL1-AS1 molecular mechanism regulating pyroptosis in HCMs, we knocked down LOXL1-AS1 and miR-761. As shown in Figure 4A, the miR-761 level was decreased by miR-761 inhibitor but markedly increased after knocking down LOXL1-AS1 in H/R-treated cells; and the knockdown of miR-761 reversed the effect of LOXL1-AS1 inhibition. The knockdown of miR-761 aggravated the decrease in cell viability and H/R treatment-induced LDH level upregulation, whereas LOXL1-AS1 knockdown showed the opposite effect; and miR-761 inhibition reversed the effects of LOXL1-AS1 knockdown (Figure 4B and C). Additionally, miR-761 knockdown reversed the inhibitory effects of knockdown LOXL1-AS1 on inflammatory factor release and pyroptosis-related protein expression in H/R-treated cells (Figure 4D and E). Moreover, miR-761 silence neutralized the inhibitory effect on pyroptosis caused by LOXL1-AS1 knockdown in H/R-treated cells (Figure 4F). In addition, H/R treatment significantly upregulated NLRP3 fluorescence intensity and LOXL1-AS1 knockdown rescued the upregulation effect of H/R treatment, whereas miR-761 knockdown reversed the effect of LOXL1-AS1 knockdown on NLRP3 (Figure 4G). Consequently, LOXL1-AS1 induced pyroptosis by decreasing the miR-761 level in H/R-treated cells.
Figure 4
LOXL1-AS1 induced human cardiomyocyte pyroptosis by targeting miR-761 under H/R conditions. (A) Using quantitative real-time polymerase chain reaction to measure the miR-761 level. (B) Using MTT to detect cell viability. (C) Using the Release Assay Kit to test the LDH level. (D) Using enzyme-linked immunosorbent assay to monitor IL-1β and IL-18 levels. (E) Using Western blot to determine ASC, cleaved caspase-1, GSDMD-N, NLRP3, IL-1β, and IL-18 levels. (F) Using flow cytometry to examine caspase-1 activity. (G) Using immunofluorescence to detect NLRP3 expression. Data were presented as mean±standard deviation, and 3 separate experiments were performed in triplicate.
GAPDH = glyceraldehyde-3-phosphate dehydrogenase; GSDMD = gasdermin D; H/R = hypoxia/reoxygenation; IL = interleukin; LDH = lactate dehydrogenase; LOXL1-AS1 = lysyl oxidase like 1 antisense RNA 1; NLRP3 = NOD-like receptor pyrin domain-containing protein 3.
*p<0.05; **p<0.01; ***p<0.001.
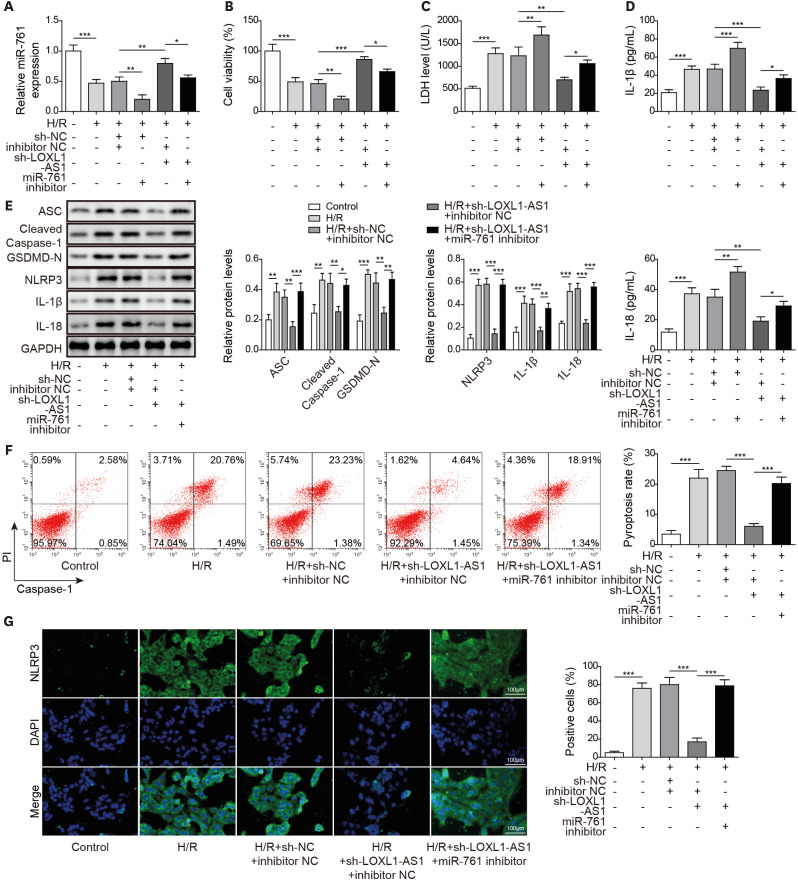
MiR-761 targeted phosphatase and tensin homolog
Using the Starbase database, we discovered that miR-761 and PTEN have mutual binding sites (Figure 5A). Further dual-luciferase verification revealed that the luciferase activity of the PTEN WT group was repressed by miR-761 mimics and promoted by miR-761 inhibitor, whereas the luciferase activity of the PTEN MUT group remained unchanged compared with that of the miR-NC group, confirming the mutual binding relationship between miR-761 and PTEN (Figure 5B). Furthermore, miR-761 overexpression significantly downregulated PTEN mRNA and protein levels, whereas miR-761 knockdown significantly upregulated PTEN mRNA and protein levels in HCMs (Figure 5C and D). Besides, following miR-761 overexpression under H/R conditions, PTEN mRNA and protein levels in HCMs were downregulated (Figure 5E and F). In general, PTEN was a downstream target of miR-761 in HCMs.
Figure 5
MiR-761 targeted PTEN. (A) Using bioinformatics to predict miR-761 and PTEN binding sites. (B) Using dual-luciferase to verify the miR-761 and PTEN mutually binding relationship. (C, D) Using qRT-PCR and Western blot to determine the PTEN level. (E, F) Using qRT-PCR and Western blot to monitor the PTEN level. Data were presented as mean±standard deviation, and 3 separate experiments were performed in triplicate.
GAPDH = glyceraldehyde-3-phosphate dehydrogenase; H/R = hypoxia/reoxygenation; MUT = mutant type; PTEN = phosphatase and tensin homolog; qRT-PCR = quantitative real-time polymerase chain reaction; WT = wild-type.
***p<0.001.
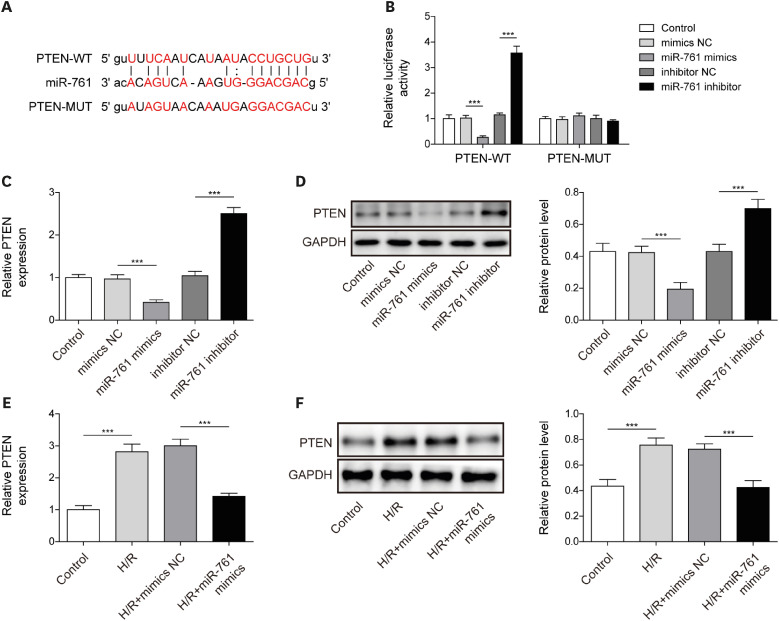
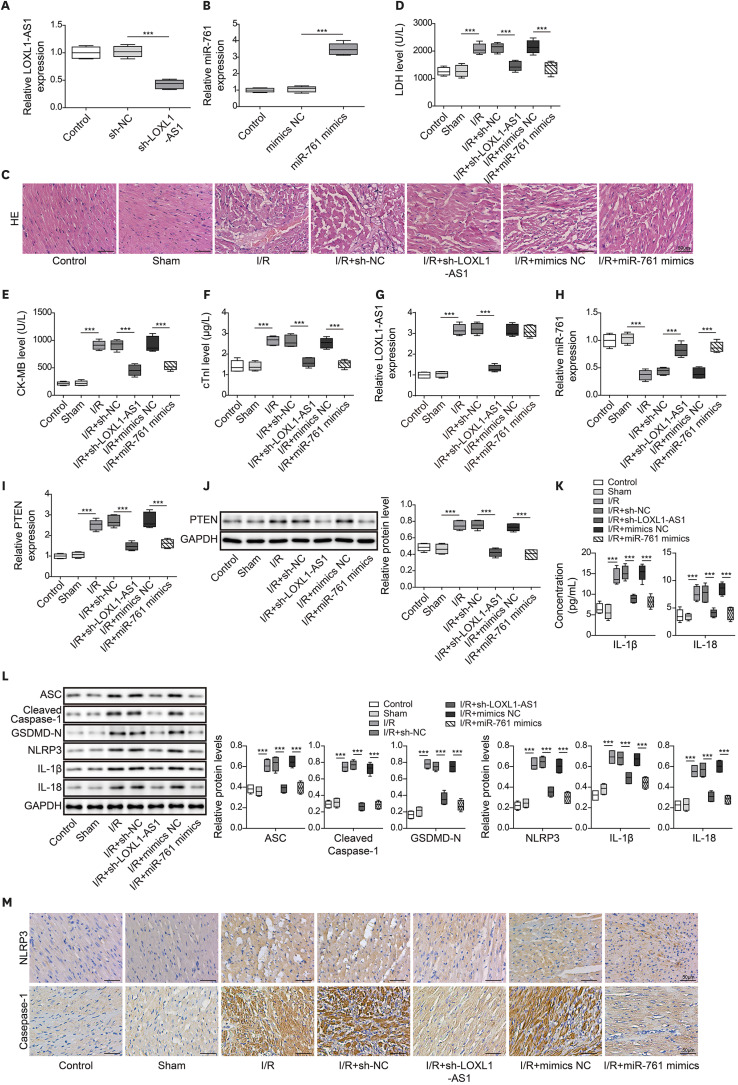
Long noncoding RNA lysyl oxidase like 1 antisense RNA 1 enhanced myocardial ischemia and reperfusion injury by regulating miR-761 in mice
Finally, we verified LOXL1-AS1/miR-761/PTEN function in I/R mice. As shown in Figure 6A and B, sh-LOXL1-AS1 significantly decreased the LOXL1-AS1 level, and miR-761 mimics upregulated the miR-761 level in myocardial tissues. Figure 6C found that the myocardial fibers of mice in the control and sham groups were neatly arranged without obvious breaks or necrotic spaces, and the myocardial nuclei were fusiform or oval. The I/R treatment caused myocardial cell injury, which manifested as myocardial fiber rupture and dissolution, increased muscle space, and reduced myocardial cells. In the I/R+sh-LOXL1-AS1 and I/R+miR-761 mimics group, myocardial fibrosis and reduction of myocardial cells were significantly improved. The detection of myocardial injury markers showed that LDH, CK-MB, and cTnI levels in the serum of I/R model mice were significantly upregulated, and LOXL1-AS1 knockdown or miR-761 overexpression inhibited LDH, CK-MB, and cTnI levels (Figure 6D-F). Additionally, LOXL1-AS1 and PTEN levels in the myocardial tissues of I/R model mice were significantly upregulated, and miR-761 levels were markedly downregulated. LOXL1-AS1 knockdown or miR-761 overexpression repressed PTEN levels and increased the miR-761 level. Moreover, LOXL1-AS1 knockdown inhibited the LOXL1-AS1 level in the myocardial tissues of I/R model mice, and miR-761 overexpression had no effect on the LOXL1-AS1 level (Figure 6G-J). The upregulated release of IL-1β and IL-18 in I/R mice was downregulated by LOXL1-AS1 knockdown or miR-761 overexpression (Figure 6K). Moreover, WB and IHC detection revealed that LOXL1-AS1 knockdown suppressed the high expression of inflammatory factors and pyroptosis-related proteins in I/R model mice (Figure 6L and M). These results demonstrated that LOXL1-AS1 enhanced mouse cardiomyocyte pyroptosis and promoted MIRI by targeting miR-761 to regulate PTEN expression.
Figure 6
LOXL1-AS1 enhanced myocardial ischemia and reperfusion injury by regulating miR-761 in mice. (A, B) Using qRT-PCR to monitor LOXL1-AS1 and miR-761 levels. (C) Using HE staining to measure changes in the myocardial tissue morphology (scale bar: 50 μm). (D-F) Using the Release Assay Kit to detect LDH, CK-MB, and cTnI levels in serum. (G, H) Using qRT-PCR to determine LOXL1-AS1 and miR-761 levels. (I, J) Using qRT-PCR and Western blot to monitor the PTEN level. (K) Using ELISA to measure IL-1β and IL-18 levels. (L) Using Western blot to examine ASC, cleaved caspase-1, GSDMD-N, NLRP3, IL-1β, and IL-18 levels. (M) Immunohistochemistry detection of NLRP3 and caspase-1 expression in the myocardial tissue (scale bar: 50 μm). Data are presented as mean±standard deviation (n=5).
CK-ME = creatine kinase-MB; cTnI = cardiac troponin I; GAPDH = glyceraldehyde-3-phosphate dehydrogenase; GSDMD = gasdermin D; HE = hematoxylin and eosin; IL = interleukin; I/R = ischemia/reperfusion; LDH = lactate dehydrogenase; LOXL1-AS1 = lysyl oxidase like 1 antisense RNA 1; NLRP3 = NOD-like receptor pyrin domain-containing protein 3; PTEN = phosphatase and tensin homolog; qRT-PCR = quantitative real-time polymerase chain reaction.
***p<0.001.
DISCUSSION
MIRI can cause adverse cardiovascular consequences after myocardial ischemia, cardiac surgery, or circulatory arrest.16) Pyroptosis is a form of inflammatory cell death, which may be driven by activating NLRP3 inflammasomes after myocardial infarction.17) However, the molecular mechanism of pyroptosis in MIRI remains largely unknown. To date, we examined the specific mechanism of pyroptosis in MIRI through in vivo and in vitro experiments, which involved LOXL1-AS1/miR-761/PTEN. We observed that LOXL1-AS1 promoted PTEN expression by targeting miR-761 to enhance cardiomyocyte pyroptosis. This is the first time we have reported the LOXL1-AS1/miR-761/PTEN study of pyroptosis in MIRI. LOXL1-AS1 and miR-761 are expected to be clinical diagnostic molecules for myocardial ischemia-reperfusion.
LncRNAs are a variety of RNAs involved in several biological processes in various branches of life. At present, numerous lncRNAs have been reported to participate in MIRI. Liang et al.18) reported that the lncRNA ROR/miR-124-3p/TRAF6 axis regulated the ischemia–reperfusion injury-induced inflammatory response in HCMs. Tan et al.19) reported that lncRNA MIAT knockdown alleviated oxygen–glucose deprivation-induced HCM injury by regulating the JAK2/STAT3 pathway via miR-181a-5p. Furthermore, lncRNA ROR knockdown alleviated H/R-induced pyroptosis and inflammation.20) Of note, LOXL1-AS1 was significantly upregulated in the serum of patients with atherosclerosis, and the LOXL1-AS1/miR-515-5p/STAT3 axis promoted atherosclerotic cell proliferation and migration.21) Furthermore, LOXL1-AS1 enhanced ox-LDL-induced pyroptosis in human coronary endothelial cells by binding miR-16-5p.9) According to the literature, in coronary heart disease, LOXL1-AS1 was noted to be significantly upregulated in the serum of patients with coronary heart disease and atherosclerosis. Silencing LOXL1-AS1 could promote HCAEC viability and reduce pyroptosis.9) The above suggests that LOXL1-AS1 is closely associated with cardiovascular disease and that it may serve as a clinically significant predictor. However, its biological effects and regulatory mechanisms have not yet been explained in MIRI. Pyroptosis is an inflammatory form of death. NLRP3 inflammasome leads to the maturation and secretion of inflammatory cytokines, including IL-1β, and induces pyroptosis through caspase-1 activation.22) Caspase-1 induces pyroptosis by cleaving GSDMD protein. The cleaved GSDMD releases its N-terminal domain, which has the activity to bind membrane lipids and punch holes in the cell membrane, thereby leading to changes in cell osmotic pressure and swelling until the eventual cell membrane rupture.23) Congruously, our results suggested that H/R treatment dramatically increased ASC, cleaved caspase-1, GSDMD-N, NLRP3, IL-1β, and IL-18 levels. Furthermore, we noted that sh-LOXL1-AS1 could improve H/R-induced HCM pyroptosis. To the best of our knowledge, this is the first study to report on the regulation role of LOXL1-AS1 on cell pyroptosis under H/R conditions.
CeRNAs mainly refer to lncRNAs or circRNAs that can competitively bind to miRNA, interfering with the binding of miRNA and its downstream mRNA, thus reducing the inhibition of miRNA on downstream genes and thereby affecting cell function. In recent decades, there is increasing evidence has shown that that the mechanism of ceRNAs is widely involved in the regulation of myocardial cell injury. For example, lncRNA XIST facilitated hypoxia-induced HCM injury by targeting miR-191-5p.24) Through bioinformatics prediction and verification, we observed that LOXL1-AS1 and miR-761 have mutual binding sites. According to the literature, miR-761 could suppress human vascular smooth muscle cell proliferation and migration by targeting bone morphogenetic protein 9.25) Additionally, miR-761 inhibited the mitochondrial fission factor by inhibiting mitochondrial division and cardiomyocyte apoptosis.15) It was previously reported that miR-761 reduced the secretion of inflammatory cytokines, IL-1β and IL-18 (pyroptosis is accompanied by IL-1β and IL-18 secretion),13) suggesting that miR-761 may improve cardiomyocyte pyroptosis. In this study, we observed that the miR-761 level was markedly downregulated following H/R treatment, and miR-761 overexpression improved H/R-induced HCM pyroptosis. We first reported that LOXL1-AS1 induced cardiomyocyte pyroptosis by inhibiting miR-761.
PTEN is a lipid and protein bisphosphatase, which can antagonize pathways, including PI3K, and regulate cell survival and growth.26) Studies have shown that the PTEN subtype, PTENα, acted as a mitochondrial quality controller to maintain mitochondrial function and cardiac homeostasis.27) PTEN inhibition plays a vital role in protecting the heart from MIRI. Cui et al.28) reported that knocking down PTEN could reduce myocardial pathological damage and inhibit NLRP3-mediated myocardial cell apoptosis. Dai et al.29) reported that miR-129 alleviated MIRI in rats by inhibiting PTEN expression. Through bioinformatics prediction and verification, we noted that miR-761 and PTEN had mutual binding sites. Moreover, we demonstrated the targeting effects between miR-761 and PTEN in cardiomyocytes. In vivo, we further verified that LOXL1-AS1 enhanced mouse cardiomyocyte pyroptosis and promoted MIRI by regulating miR-761/PTEN axis.
In summary, we explored the LOXL1-AS1 mechanism in cardiomyocyte pyroptosis in MIRI. We have revealed through in vivo and in vitro experiments that LOXL1-AS1 enhanced cardiomyocyte pyroptosis and promoted MIRI by targeting miR-761 to increase PTEN expression. We observed that sh-LOXL1-AS1 or miR-761 mimics ameliorated H/R-induced cardiomyocyte pyroptosis. By specifically interfering with the LOXL1-AS1 and miR-761 levels in cardiomyocytes, it is of great significance to directly reflect the role of LOXL1-AS1 and miR-761 in MIRI, thereby providing new ideas for the study of molecular mechanism and clinical treatment of MIRI.
 XML Download
XML Download