

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Regular exercise enhances endurance and decreases the risk of several chronic diseases, such as type 2 diabetes, obesity, hypertension, coronary heart disease, sarcopenia, and cardiovascular diseases [1]. Physical exercise increases energy maintenance through mitochondrial biogenesis, mitochondrial oxidative capacity, and improvements in the skeletal muscle function [2]. Exercise promotes the consumption of energy sources, such as glycogen and glucose. Glucose transport is increased by muscle contractions, maximizing the internal energy metabolism. On the other hand, hard work or intensive exercise can cause active free radical production, resulting in tissue oxidative damage. Physical fatigue is caused mainly by the depletion of energy and the excessive accumulation of metabolites.
Recently, the development of natural supplements with efficient recovery ability after exercise, which accelerates the elimination of fatigue-related metabolites, is becoming a major research focus. Gynostemma pentaphyllum (Thunb.) Makino is an herbaceous vine plant of the family Cucurbitaceae, whose leaves are used for traditional tea or oriental herb medicine in Asia [3]. G. pentaphyllum has been prescribed traditionally for clearing heat, detoxification, dry cough, and chronic bronchitis [4]. Several studies have reported that G. pentaphyllum has important biological activities, including anti-oxidant [5], anti-inflammatory [678], hypoglycemic [9], hypolipidemic [101112], anti-obesity [13], anti-diabetic [14], neuroprotective [15], and anti-cancer properties [16]. G. pentaphyllum contains saponins, polysaccharides, flavonoids, and other chemicals [1718]. Gypenosides are major active components of dammarane-type saponins extracted from G. pentaphyllum. Gypenosides have anti-oxidant [19], anti-cancer [20], hypolipidemic [10], hepatoprotective [1019], and anti-obesity activities [13]. The no observed adverse effect level of the G. pentaphyllum oral extract in rats was reported to be 750 mg/kg [21]. Recent studies have shown that gypenosides and polysaccharides from G. pentaphyllum extend the swimming exhaustion time and increase the hepatic and muscle glycogen concentrations in mice [34222324]. Although the beneficial effects of G. pentaphyllum have been proven, the possible energy metabolism and associated molecular mechanisms behind the ability of the G. pentaphyllum extract (GPE) to increase the exercise capacity of mice on treadmill exercise is unknown.
Recently, a beneficial method that produces GPE with much higher contents of gypenoside L (GL), gypenoside LI (GLI), and ginsenoside Rg3 (Rg3) than the extracts by other conventional methods was developed [25]. The previous study reported that treatment with GPE and GL increased the glucose uptake and glucose transporter type 4 expression by activating the adenosine monophosphate (AMP)-activated protein kinase (AMPK) and acetyl-CoA carboxylase signaling pathway, which enhances the mRNA expression of peroxisome proliferator-activated receptor-γ coactivator-1α (PGC-1α) in L6 skeletal muscle. In addition, GPE reduced the body fat mass and increased the lean body mass by AMPK activation. GPE supplementation inhibited adipogenesis in high-fat diet induced obese C57BL/6 mice [13]. PGC-1α, a transcription factor coactivator, plays an important role in regulating the genes involved in mitochondrial biogenesis, glucose/fatty acid metabolism, angiogenesis, oxidative metabolism, fiber-type switching, and muscle growth. Exercise drastically increased PGC-1α expression in the muscles. The overexpression of PGC-1α in mice improved the exercise capacity, oxygen uptake, and fatigue resistance [25]. Hence, effective gypenosides, GPE and GL, improved exercise performance by activating PGC-1α. Thus, this study examined the effects of GPE and GL on exercise performance and energy metabolism-related gene expression in treadmill-trained mice.
MATERIALS AND METHODS
Materials
G. pentaphyllum was purchased from Hunan Tea Group Co., Ltd. (Changsha, China). Professor Zhou Ribao was responsible for identifying the plant species. A voucher specimen was deposited at the College of Medicine, Hunan University of Traditional Chinese Medicine (HUCM) Herbarium under number 20210301-1. GPE and GL were obtained from BTC Corporation (Ansan, Korea) [2627]. Briefly, the dried leaves of G. pentaphyllum were extracted with hot water and a 50% aqueous ethanol solution. The resulting extracts were filtered and evaporated to dryness. The GL content in GPE was analyzed by high-performance liquid chromatography [2627]. The developed GPE contained much higher GL, GLI, and Rg3 contents than the products from conventional extraction methods (18 mg, 14 mg, and 1.5 mg, respectively, per g of G. pentaphyllum leaf extract). Creatine monohydrate (CrM) (Sigma-Aldrich Inc., St. Louis, MO, USA) was used as the positive control.
Animals and experimental design
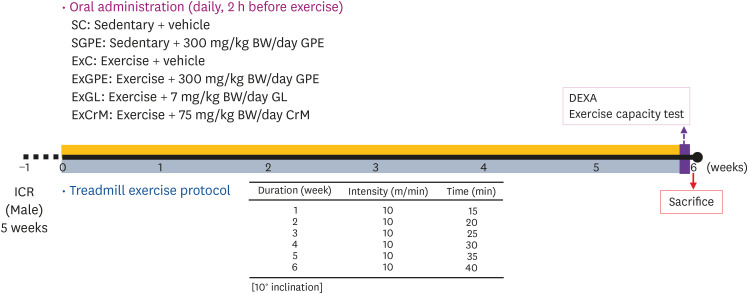
The Animal Care and Use Committee of Hallym University approved this study (Hallym 2019-28). Male ICR mice (5 weeks) were supplied by Doo Yeol Biotech Co., Ltd. (Seoul, Korea) and fed a standard laboratory diet (Cargill Agri Purina, Inc., Seongnam, Korea) and with access to tap water ad libitum. The mice were acclimatized for 1 week before the experiments and housed under a constant room temperature (23 ± 3°C) and humidity (50 ± 10%) controlled room with a 12 h light/dark cycle (lights on from 8:00 AM to 8:00 PM). A 2-way experiment (Exercise X Treatment) was designed for the 6 groups (n = 10 per group in each test): sedentary + vehicle (SC), sedentary + 300 mg/kg body weight (BW)/day GPE (SGPE), exercise + vehicle (ExC), exercise + 300 mg/kg BW/day GPE (ExGPE), exercise + 7 mg/kg BW/day GL (ExGL), and exercise + 75 mg/kg BW/day creatine monohydrate (ExCrM). GPE, GL, and CrM were dissolved in a saline solution and administered daily by oral gavage for 6 weeks. Treadmill training was conducted using a single-lane rodent treadmill (Exer-3R Treadmill; Columbus Instruments, Columbus, OH, USA) supplied with a shocking grid at the rear. The mice were selected based on their exercise compliance and subjected to weekly training sessions involving running speeds of 10 m/min over a 15-min period with a 10° inclination to the treadmill environment acclimatization. The exercise duration was increased gradually from 15 to 40 min per week (Fig. 1). The bodyweight, dietary, and social behaviors were monitored during the administration period and exercise.
Fig. 1
Experimental design to examine the effects of GPE, GL, CrM supplementation on exercise adaptation. The animals were assigned randomly to the indicated 6 groups (n = 10 per group in each test). Physical exercise capacity and related assessments were conducted during the test for 6 weeks.
GPE, G. pentaphyllum extract; GL, gypenoside L; BW, body weight; CrM, creatine monohydrate; SC, sedentary with vehicle; SGPE, sedentary with 300 mg/kg BW/day of GPE; ExC, exercise with vehicle; ExGPE, exercise with 300 mg/kg BW/day of GPE; ExGL, exercise with 7 mg/kg BW/day of GL; ExCrM, exercise with 75 mg/kg BW/day of CrM.
Exercise endurance test
An exercise endurance test was conducted after 6 weeks of GPE, GL, and CrM administration and exercise. The exercise capacity was determined by increasing the treadmill speed from a 10 m/min period with 10° inclination in 1 m increments every 1 min until exhaustion (> 25 m/min maximum speed). The point of exhaustion was defined as when mice remained in continuous contact with the shocking grid for 10 s. The exercise capacity was expressed as the work done in Joules (kg m2s−2).
Body composition analysis and sample collection
The whole-body composition (fat mass and lean body mass percentage) was measured using dual-energy X-ray absorptiometry (DEXA, PIXlmusTM; GE Lunar, Madison, WI, USA) before one day the end of the experiment. After 16 h fasting, the mice were then anesthetized using tribromoethanol diluted with tertiary amyl alcohol. Blood samples were taken from the orbital venous plexus of the mice using capillary tubes. Blood was then centrifuged at 5,000 rpm for 10 min to separate the serum, which was stored at −70°C until further analysis. The visceral organs, including the quadriceps femoris muscle (QF), gastrocnemius muscle (GA), SOL, extensor digitorum longus muscle (EDL), and liver were excised and weighed accurately after sacrifice via a cervical dislocation. The organs were kept at −70°C until further analysis.
Blood chemistry assay
Serum levels of glucose, triglyceride (TG), total cholesterol (CHOL), low-density lipoprotein-cholesterol (LDL-CHOL), high-density lipoprotein-cholesterol (HDL-CHOL), blood urea nitrogen (BUN), and creatinine (CREA), as well as the serum activities of lactate dehydrogenase (LDH), alanine aminotransferase (ALT), aspartate aminotransferase (AST), and alkaline phosphatase (ALP) were measured using a blood chemistry autoanalyzer (KoneLab 20XT, Thermo Fisher Scientific, Vantaa, Finland). The serum lactate levels were analyzed using commercial enzyme-linked immunosorbent assay kits (Abcam, Cambridge, MA, USA) according to the manufacturer's instructions.
Tissue glycogen content assay
For each mouse, 100 mg of liver and muscle (GA) was weighed, transferred to a tube, and homogenized in 1 mL of ice-cold phosphate-buffered saline. After centrifugation at 12,000 rpm for 10 min at 4°C, the supernatant was decanted and kept on ice. The glycogen content in the liver and muscle was analyzed using a commercial kit (Abcam), according to the manufacturer's instructions.
Protein expression analysis
The muscle (GA) was homogenized in ice-cold lysis buffer (20 mM HEPES, pH 7.5, 150 mM NaCl, 1% Triton X-100, 1 mM EDTA, 1 mM EGTA, 100 mM NaF, 10 mM sodium pyrophosphate, 1 mM sodium orthovanadate, 20 μg/mL aprotinin, 10 μg/mL anti-pain, 10 μg/mL leupeptin, 80 μg/mL benzamidine HCl, and 0.2 mM phenylethylsulfonyl fluoride). The insoluble material was removed by centrifugation at 12,000 rpm for 10 min, and the supernatant was collected for western blot analysis. The protein content of the lysates was measured using a BCA protein assay kit (Thermo Scientific, Rockford, IL, USA). The proteins (50 μg) were separated by 10% SDS-PAGE and transferred to a polyvinylidene difluoride membrane (Millipore, Billerica, MA, USA). The membranes were blocked for 1 h in 5% skim milk-TBST (20 mM Tris-HCl, pH 7.5, 150 mM NaCl, 0.1% Tween 20) at room temperature and then incubated overnight at 4°C with anti-phospho-AMPKα (Thr172), anti-AMPKα, anti-phospho-p38 (Thr180/Tyr182), anti-p38, anti-PGC-1α, anti-β-actin (Cell Signal Technology, Beverly, MA, USA), anti-phospho-PGC-1α (R&D system Inc., Minneapolis, MN, USA), anti-NRF2, and anti-phospho-NRF2 (Ser40) (Abcam) primary antibodies. Subsequently, the membranes were incubated with horseradish peroxidase-conjugated goat anti-rabbit or mouse IgG. The immunoreactive bands were detected with Luminata TM Forte Western HRP Substrate (Millipore) and quantified using an ImageQuantTM LAS 500 imaging systems (GE Healthcare Bio-Sciences AB, Uppsala, Sweden). β-actin was used as the internal control.
Real-time reverse-transcription polymerase chain reaction (RT-PCR)
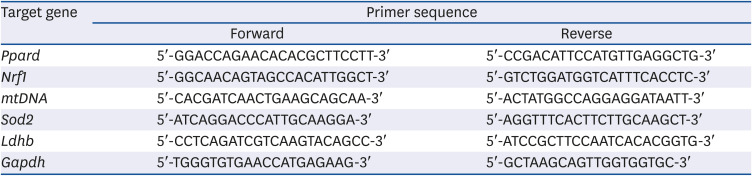
The total RNA from the skeletal muscle (SOL) was extracted using TRIzol reagent (Roche, Indianapolis, IN, USA). cDNA was synthesized from 2 μg of the total RNA using a HyperScriptTM RT master mix kit (GeneAll Biotechnology, Seoul, Korea). Real-time PCR for various genes was performed using Rotor-gene 300 PCR (Corbett Research, Mortlake, NSW, Australia) and Rotor-GeneTM SYBR Green Kit (QIAGEN, Hilden, Germany). The data were then analyzed using the Rotor-gene 6000 series System Software program (Corbett Research), and the values were normalized to those of Gapdh. Table 1 lists the various primers used for real-time PCR analysis.
Table 1
Quantitative real-time polymerase chain reaction primers
RESULTS
Effects of GPE administration on the body and muscle weight in treadmill-trained mice
The effects of GPE supplementation on exercise performance were evaluated by administering GPE, GL, and CrM, or saline solution (SC and ExC) orally to the mice. Table 2 lists the bodyweight, fat mass, lean mass, and muscle weight. The weights of the experimental groups were monitored continuously during the 6 weeks of exercise, supplementation, or both. At the beginning of the experiment, there was no significant difference in the initial BWs among the experimental groups. The supplementation of GPE without exercise for 6 weeks induced a significant decrease in BW and fat mass, but the lean mass percentage was significantly higher in the SGPE group than the SC group. In the ExC group, significant changes were observed in the final BW gain, fat mass, and lean mass percentage, compared to the SC group. In addition, the ExGL group showed a significant decrease in BW gain, while the ExGPE group showed no significant differences in BW gain. Significant differences in the fat mass percentage among the groups were observed, which was significantly decreased compared to the ExC group, while the lean mass percentage of ExGPE showed the greatest relative increase, 84.6 ± 0.8%, which was higher than that in the ExC group. There were no significant differences in the BW, fat mass, and lean mass in the ExCrM group. In terms of muscle mass, SGPE increased the SOL and EDL muscle weight significantly compared to the SC group but did not change the QF and GA muscle weight. Moreover, the muscle masses of GA, SOL, and EDL in the ExC group were increased by 0.104, 0.013, and 0.016 g/100 g BW, respectively, compared to the SC group. No significant differences between the ExGPE and ExCrM groups were observed relative to the ExC group. On the other hand, the GA muscle weights were significantly higher in the ExGL group than the ExC group, but there were no changes in the QF, SOL, and EDL groups.
Table 2
Effect of GPE and GL treatment on BW, fat mass percentage, lean mass percentage, and QF, GA, SOL, and EDL weights in treadmill-trained ICR mice
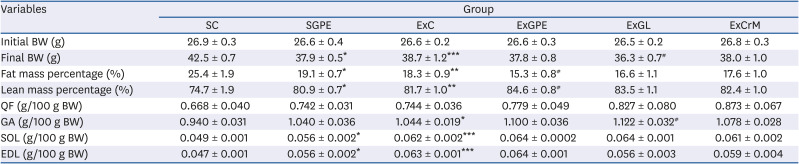
Values are the means ± SEM for n = 10 per group.
GPE, G. pentaphyllum extract; GL, gypenoside L; QF, quadriceps femoris muscle; GA, gastrocnemius muscle; SOL, soleus muscle; EDL, extensor digitorum longus muscle; BW, body weight; CrM, creatine monohydrate; SC, sedentary with vehicle; SGPE, sedentary with 300 mg/kg BW/day of GPE; ExC, exercise with vehicle; ExGPE, exercise with 300 mg/kg BW/day of GPE; ExGL, exercise with 7 mg/kg BW/day of GL; ExCrM, exercise with 75 mg/kg BW/day of CrM. *P < 0.05, **P < 0.01, and ***P < 0.001 (SC group vs. SGPE or ExC group); #P < 0.05 (ExC group vs. ExGPE, ExGL or ExCrM group).
Effect of GPE administration on the endurance exercise performance in the treadmill-trained mice
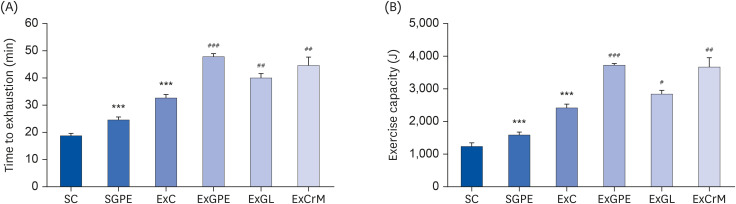
The anti-fatigue activity of GPE was determined as the endurance exercise capacity of the mice using a treadmill until exhaustion. As shown in Fig. 2A, the SGPE and ExC groups took a significantly longer time to reach exhaustion (1.31-fold and 1.74-fold, respectively) than the SC group. In addition, the ExGPE, ExGL, and ExCrM mice exercised for a longer period than those in the ExC group, demonstrating significant differences (1.46-fold with ExGPE, 1.22-fold with ExGL, and 1.37-fold with ExCrM) compared to the ExC group. GPE, GL, and CrM supplementation were associated with significantly improved exercise capacity in the ExGPE, ExGL, and ExCrM mice, respectively, compared to the vehicle treatment (Fig. 2B). These results suggest that GPE and GL supplementation enhances endurance exercise capacity.
Fig. 2
Effects of GPE and GL treatment on treadmill exercise performance. (A) Endurance time to exhaustion and (B) exercise capacity. Values are means ± SEM for n = 10 per group. GPE, G. pentaphyllum extract; GL, gypenoside L; BW, body weight; CrM, creatine monohydrate; SC, sedentary with vehicle; SGPE, sedentary with 300 mg/kg BW/day of GPE; ExC, exercise with vehicle; ExGPE, exercise with 300 mg/kg BW/day of GPE; ExGL, exercise with 7 mg/kg BW/day of GL; ExCrM, exercise with 75 mg/kg BW/day of CrM.
***P < 0.001 (SC group vs. SGPE or ExC group); #P < 0.05, ##P < 0.01, ###P < 0.001 (ExC group vs. ExGPE, ExGL or ExCrM group).
Effect of GPE administration on fatigue-associated biochemistry
The serum collected 30 min after the exercise test at 6 weeks was analyzed to investigate the effects of GPE and GL on the anti-fatigue-associated indices. During intense exercise, the energy supply comes from glycogen degradation by phosphorylase and circulating glucose released by the liver. The glucose levels in serum are an important marker for exercise performance maintenance [2829]. As shown in Table 3, the serum glucose levels in the ExGPE, ExGL, and ExCrM groups were 25.1%, 38.2%, and 57.9%, respectively, compared to the ExC group. The serum TG levels were also increased significantly in the ExGPE group compared to that in the ExC group. No significant differences were observed between the groups with respect to the other indices, such as CHOL, LDL-CHOL, or HDL-CHOL levels, compared to those in the ExC groups. Intensive exercise increased the creatine kinase (CK) and LDH markers, indicating muscle damage. As shown in Table 3, the CK levels in the ExGPE group were significantly lower than those in the ExC group. The serum LDH levels were significantly higher in the ExC group. Although there was no significant difference in the LDH levels between the ExGPE group and the ExC group, the LDH levels of the ExGL and ExCrM groups were significantly lower. Compared to the ExC group, the serum lactate levels were 22.2%, 18.3%, and 30.98% lower in the ExGPE, ExGL, and ExCrM groups, respectively. No significant changes in the liver (ALT, AST, and ALP) and renal (BUN and CREA) profiles were observed in any group except in the ExGL group for BUN. Trend analysis showed that GPE and GL treatment had a remarkable effect on the blood glucose and TG levels and decreased the serum CK, LDH, and lactate levels.
Table 3
Effects of GPE and GL treatment on clinical biochemistry tests in ICR mice
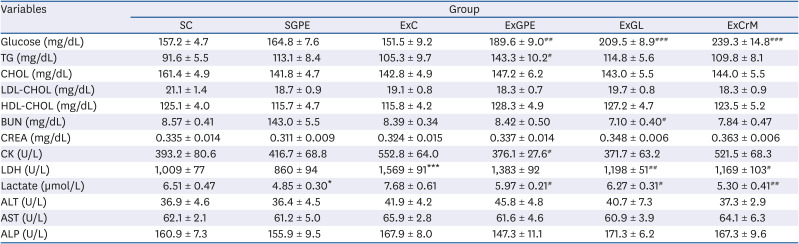
Values are the means ± SEM for n = 10 per group.
GPE, G. pentaphyllum extract; GL, gypenoside L; BW, body weight; CrM, creatine monohydrate; SC, sedentary with vehicle; SGPE, sedentary with 300 mg/kg BW/day of GPE; ExC, exercise with vehicle; ExGPE, exercise with 300 mg/kg BW/day of GPE; ExGL, exercise with 7 mg/kg BW/day of GL; ExCrM, exercise with 75 mg/kg BW/day of CrM; TG, triglyceride; CHOL, total cholesterol; LDL-CHOL, low-density lipoprotein-cholesterol; HDL-CHOL, high-density lipoprotein-cholesterol; BUN, blood urea nitrogen; CREA, creatinine; CK, creatine kinase; LDH, lactate dehydrogenase; ALT, alanine aminotransferase; AST, aspartate aminotransferase; ALP, alkaline phosphatase.
*P < 0.05, and ***P < 0.001 (SC group vs. SGPE or ExC group); #P < 0.05, ##P < 0.01, and ###P < 0.001 (ExC group vs. ExGPE, ExGL or ExCrM group).
Effect of GPE administration on the liver and muscle glycogen level
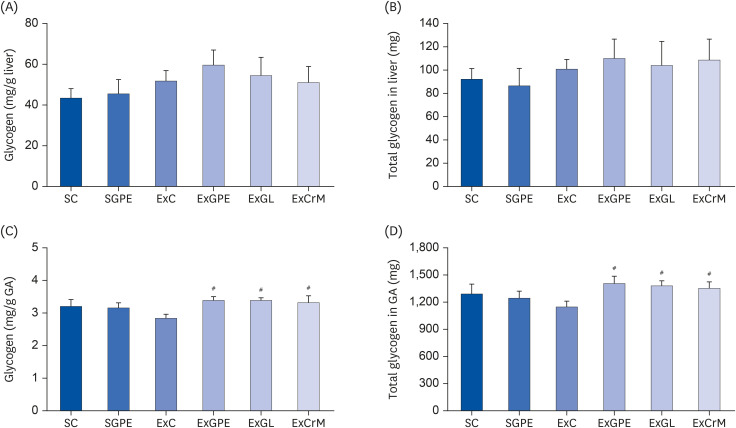
The glycogen level of liver and muscle (GA) was monitored to determine the effects of GPE supplementation on the glycogen status. As shown in Fig. 3, the liver glycogen content did not differ significantly among the groups (Fig. 3A and B). On the other hand, muscle glycogen levels exhibited a significant difference among the groups. The muscle glycogen of the ExGPE, ExGL, and ExCrM groups was significantly higher than that in the ExC group (Fig. 3C and D). These results suggest that GPE and GL supplementation during the exercise increased the muscle glycogen storage in mice and maintained a hepatic glycogen content in the mice, leading to enhanced energy reserves and blood glucose maintenance. This may be one of the mechanisms of anti-fatigue effects on GPE and GL supplementation.
Fig. 3
Effects of GPE and GL treatment on glycogen contents in the liver and muscle of ICR mice. (A) Glycogen content (mg/g liver). (B) Total glycogen in liver (mg). (C) Glycogen content (mg/g GA). (D) Total glycogen in GA (mg). The values are the means ± SEM for n = 10 per group.
GPE, G. pentaphyllum extract; GL, gypenoside L; BW, body weight; CrM, creatine monohydrate; SC, sedentary with vehicle; SGPE, sedentary with 300 mg/kg BW/day of GPE; ExC, exercise with vehicle; ExGPE, exercise with 300 mg/kg BW/day of GPE; ExGL, exercise with 7 mg/kg BW/day of GL; ExCrM, exercise with 75 mg/kg BW/day of CrM; GA, gastrocnemius muscle.
#P < 0.05 (ExC group vs. ExGPE, ExGL or ExCrM group).
Effect of GPE administration on mitochondrial biogenesis
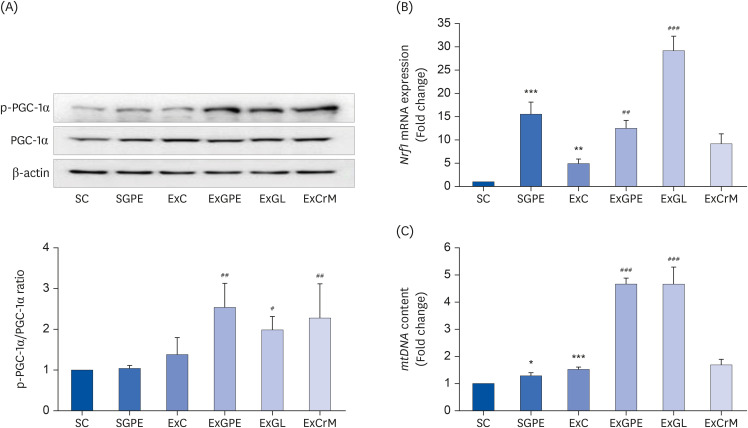
The muscles (GA and SOL) of the mice, which are used extensively during treadmill training, were collected to verify the effects of GPE supplementation on mitochondrial biogenesis in mice. The gene expression of the mitochondrial biogenesis transcription factors and mitochondrial DNA (mtDNA) content in the muscle tissue of administrated mice was evaluated. GPE, GL, and CrM administration greatly increased the protein expression of phosphorylated PGC-1α by 1.84-fold, 1.42-fold, and 1.65-fold, respectively, compared to the ExC group (Fig. 4A). The nuclear respiratory factor 1 (Nrf1) gene for the expression of the key mitochondrial genes was also upregulated remarkably in the SGPE and ExC groups by 15.52-fold and 4.92-fold, respectively, compared to that of the SC group. By contrast, the levels of the Nrf1 gene in the ExGPE and ExGL groups were increased 2.54-fold and 5.93-fold compared to the ExC group (Fig. 4B). On the other hand, ExCrM mice did not show any detectable changes in Nrf1 gene expression compared to the ExC group. The mtDNA content also increased markedly in the SGPE group and ExC group by 1.29-fold and 1.52-fold, respectively, compared to that of the SC group. In particular, both the ExGPE group and ExGL group were 3.06-fold higher than that in the ExC group (Fig. 4C). The ExCrM group did not affect the mtDNA content. These results suggest that the GPE and GL treatments elevated mitochondrial biogenesis by promoting PGC-1α expression.
Fig. 4
Effects of the GPE and GL treatment on the protein and mRNA expression of mitochondrial biogenesis genes in the soleus muscle of ICR mice. (A) Protein levels of phosphorylated PGC-1α and PGC-1α were analyzed using a western blot assay. β-actin served as an internal control. The ratio of p-PGC-1α/PGC-1α was determined (n = 4 per group). The mRNA levels of Nrf1 (B), and mtDNA content (C) were analyzed by qRT-PCR. The target mRNA expression was normalized to Gapdh. The values are means ± SEM for n = 8–10 per group.
GPE, G. pentaphyllum extract; GL, gypenoside L; BW, body weight; CrM, creatine monohydrate; SC, sedentary with vehicle; SGPE, sedentary with 300 mg/kg BW of GPE; ExC, exercise with vehicle; ExGPE, exercise with 300 mg/kg BW/day of GPE; ExGL, exercise with 7 mg/kg BW/day of GL; ExCrM, exercise with 75 mg/kg BW/day of CrM; p-PGC-1α, phosphorylated peroxisome proliferator-activated receptor γ coactivator 1-alpha; PGC-1α, peroxisome proliferator-activated receptor γ coactivator 1-alpha; Nrf1, nuclear respiratory factor 1; mtDNA, mitochondrial DNA; qRT-PCR, quantitative reverse-transcription polymerase chain reaction; Gapdh, glyceraldehyde 3-phosphate dehydrogenase.
*P < 0.05, **P < 0.01, ***P < 0.001 (SC group vs. SGPE or ExC group); #P < 0.05, ##P < 0.01, ###P < 0.001 (ExC group vs. ExGPE, ExGL, or ExCrM group).
Effect of GPE administration on PGC-1α targeted genes expression
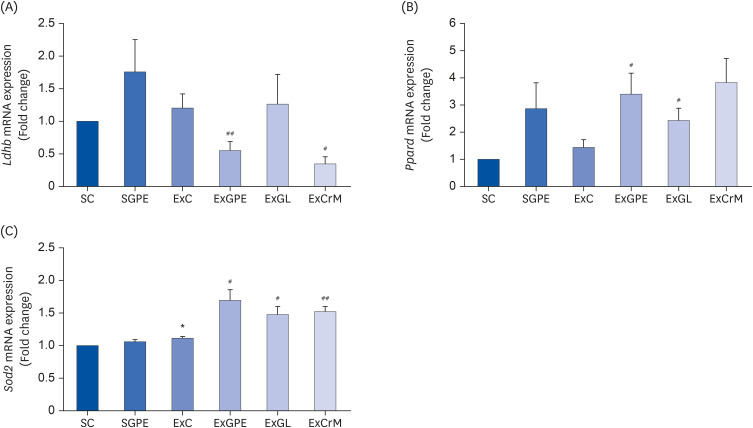
In skeletal muscles, PGC-1α acts as a transcriptional coactivator of several exercise-related genes responsible for processes, such as lactate metabolism, fatty acid metabolism, and antioxidant response [3031]. The mRNA expression of PGC-1α-related genes, including LDH B, peroxisome proliferator-activated receptor-δ (PPARδ), and superoxide dismutase 2 (SOD2) in the muscle (SOL) were measured to investigate the molecular basis of GPE for the exercise performance effect. As shown in Fig. 5A, the SGPE and ExC group increased the mRNA expression level of Ldhb, even though there was no significant difference in Ldhb mRNA expression between the SC groups. In contrast, the Ldhb mRNA level showed a trend toward a significant decrease in the ExGPE and ExCrM groups compared to that in the ExC group. The mRNA expression of Ppard was upregulated in the muscles of the ExGPE and ExGL group compared to that in the ExC group (Fig. 5B). On the other hand, the ExCrM group showed no significant activity. The ExGPE, ExGL, and ExCrM groups also showed significantly upregulated mRNA expression of Sod2 in the muscle compared to the ExC group (Fig. 5C).
Fig. 5
Effects of the GPE and GL treatment on the mRNA expression of PGC-1α targeted genes in the soleus muscle of ICR mice. The mRNA levels of Ldhb (A), Ppard (B), and Sod2 (C) were analyzed by qRT-PCR. The target mRNA expression was normalized to Gapdh. Values are means ± SEM for n = 4–6 per group.
GPE, G. pentaphyllum extract; GL, gypenoside L; BW, body weight; CrM, creatine monohydrate; SC, sedentary with vehicle; SGPE, sedentary with 300 mg/kg BW/day of GPE; ExC, exercise with vehicle; ExGPE, exercise with 300 mg/kg BW/day of GPE; ExGL, exercise with 7 mg/kg BW/day of GL; ExCrM, exercise with 75 mg/kg BW/day of CrM; PGC-1α, peroxisome proliferator-activated receptor γ coactivator 1-alpha; qRT-PCR, quantitative reverse-transcription polymerase chain reaction; Ldhb, lactate dehydrogenase B; Ppard, peroxisome proliferator-activated receptor-δ; Sod2, superoxide dismutase 2; Gapdh, glyceraldehyde 3-phosphate dehydrogenase.
*P < 0.05 (SC group vs. SGPE or ExC group); #P < 0.05, ##P < 0.01 (ExC group vs. ExGPE, ExGL, or ExCrM group).
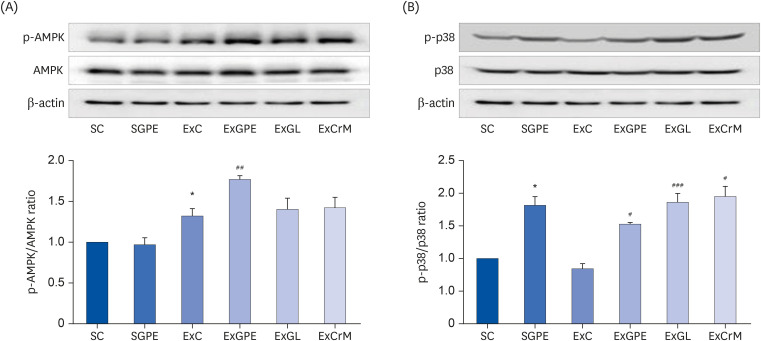
Effect of GPE administration on PGC-1α activation through AMPK and p38 phosphorylation
PGC-1α is induced by multiple signaling pathways typically activated by mitochondrial biogenesis during endurance exercise. These include the activation of AMPK, p38 MAP kinase (p38), Ca2+ mediated pathways, and reactive oxygen species (ROS)-induced signaling [323334]. Therefore, this study examined whether AMPK and p38 mediated the effects of GPE on PGC-1α activation. As shown in Fig. 6A, the ExGPE group showed 1.33 times higher AMPK phosphorylation than the ExC group. The phosphorylated p38 level was 1.81 times higher in the SGPE group than the SC group. In addition, the ExGPE, ExGL, and ExCrM groups also showed 1.8, 2.21, and 2.29 times higher levels of phosphorylated p38, respectively, than the ExC group (Fig. 6B). These results suggest that GPE and GL supplementation increased the expression of proteins involved in activating PGC-1α through AMPK and p38 phosphorylation.
Fig. 6
Effects of the GPE and GL treatment on PGC-1α activation through AMPK/p38 phosphorylation in soleus muscle of ICR mice. The protein levels of phosphorylated AMPK and AMPK (A) and phosphorylated p38, and p38 (B) were analyzed by western blot assay. β-actin served as an internal control. The ratio of p-AMPK/AMPK and p-p38/p38 were determined. The values are the means ± SEM for n = 4 per group.
GPE, G. pentaphyllum extract; GL, gypenoside L; BW, body weight; CrM, creatine monohydrate; SC, sedentary with vehicle; SGPE, sedentary with 300 mg/kg BW/day of GPE; ExC, exercise with vehicle; ExGPE, exercise with 300 mg/kg BW/day of GPE; ExGL, exercise with 7 mg/kg BW/day of GL; ExCrM, exercise with 75 mg/kg BW/day of CrM; PGC-1α, peroxisome proliferator-activated receptor γ coactivator 1-alpha; p-AMPK, phosphorylated adenosine monophosphate-activated protein kinase; AMPK, adenosine monophosphate-activated protein kinase; p-p38, phosphorylated p38 MAP kinase; p38, p38 MAP kinase.
*P < 0.05 (SC group vs. SGPE or ExC group); #P < 0.05, ##P < 0.01, ###P < 0.001 (ExC group vs. ExGPE, ExGL, or ExCrM group).
DISCUSSION
Exercise can reduce the risk of cardiovascular, metabolic, muscle, bone, joint diseases, and cancer. On the other hand, acute exercise can accelerate muscular fatigue and decrease the exercise capacity, causing muscle damage, inflammation, and oxidative stress. Muscle contractions and energy deficiency under hypoxic conditions activate the ATPases, promote glycolysis, and increase the intracellular metabolites, such as H+, lactate, Pi, and ROS. Therefore, several studies have examined the effects of reducing oxidative damage and physical fatigue and restoring the mitochondrial function through nutritional interventions, such as the consumption of synthetic products (i.e., amphetamine and caffeine) or natural dietary supplements (vitamins, minerals, and creatine) [35]. The present study revealed the role of GPE supplementation on exercise performance, clinical biochemistry, energy substrate-associated biomarkers, and mitochondrial biogenesis-related genes. The major finding of the present study was that the PGC-1α upregulation in treadmill-trained mice with GPE enhances exercise performance. From the perspective of nutritional ergogenic aids, GPE and GL are potent nutritional supplements for enhancing exercise performance and anti-fatigue effects.
Lactate in the blood is a metabolite produced during intense exercise with an insufficient oxygen supply. The blood lactate levels reflect the balance between lactate production and clearance. Hydrogen ions (H+) separate from lactate and accumulate in the muscles, which causes fatigue and depresses the muscle function and exercise performance [36]. GPE and GL supplementation and the training program effectively improved the aerobic endurance over the 6-week intervention, demonstrating a significant synergistic increase in exercise performance and exercise capacity (Fig. 2). The anti-fatigue effect of GPE and GL were reflected by the decrease in the serum CK and lactate levels and increased glucose and TG levels in mice (Table 3). GPE could facilitate the conversion of TG into fatty acids and the energy supply, resulting in a significant increase in the exercise time of GPE administration. These results suggest that muscle glycogen increased with GPE and GL supplementation in terms of the glycogen storage capacity, suggesting that improved muscle glycogen storage led to the main beneficial effect for extended treadmill exercise time to exhaustion (Fig. 3).
Exercise requires energy expenditure by ATP-generating metabolic pathways, including glucose utilization, liver glycogenolysis, lipid metabolism, and the TCA cycle [37]. During intense exercise, the energy supply initially comes from glycogen degradation by phosphorylase and circulating glucose released by the liver. The depletion of liver glycogen may lead to hypoglycemia, impaired nervous function in over-exercise fatigue conditions [38]. TG represents a critical energy source for endurance exercises. The use of adipose and TG during exercise depends on the degree of adaptability and exercise intensity. Carbohydrates and lipids are the major energy sources in the first 30 min of mild- or moderate-intensity exercise (25–65% of VO2 max) [3940]. The uptake of free fatty acids from plasma into the muscles gradually increases during 1–4 h of prolonged exercise, which is associated with increased fat oxidation because of the increased muscular energy requirements. High-intensity exercise increases the acetyl-CoA levels derived from glycogen by muscle glycogenolysis. Therefore, the blood TG and glucose or glycogen levels in the muscle and liver are important indices of maintenance performance, which can delay physical fatigue [41]. These effects of G. pentaphyllum on enhanced exercise performance have been reported with polysaccharides and gypenosides [3424]. Wang et al. [3] reported that G. pentaphyllum polysaccharide supplementation could increase the swimming exhaustion time and liver glycogen content and reduce oxidative stress. Lin-Na et al. [4] reported that the treatment with polysaccharides from G. pentaphyllum prolonged the swimming exhaustion time of the rats and enhanced the concentration of hepatic and muscular glycogen in rats. Ding et al. [24] reported that gypenosides from G. pentaphyllum showed a significantly longer swimming time to exhaustion, lower blood lactate levels, and higher blood glucose concentrations than in the control group. This study showed that GPE and GL enhanced the exercise exhaustion time and serum glucose and TG levels and improved muscle glycogen. These observations suggest that GPE and GL may enhance the endurance capacity and exert anti-fatigue effects.
PGC-1α controls the expression of multiple transcription factors associated with the lipid metabolism, metabolic homeostasis, fat browning, mitochondrial biogenesis, energy metabolism, anti-oxidative stress, and fiber-type switching. Moreover, its activity can be regulated by AMPK phosphorylation and p38 [3042]. In addition, PGC-1α activity is controlled by sirtuin-1, which is an NAD+-dependent deacetylase. PGC-1α also modulates the expression of nuclear and mitochondrial genes that are activated by NRF1and NRF2 and the estrogen-related receptor alpha transcription factor. The increased expression of transcription factor A mitochondria controls mtDNA replication and transcription [4344]. These results showed that GPE and GL increased the expression of PGC-1α phosphorylation significantly and enhanced Nrf1 and mtDNA expression, which was higher than the CrM treatment (Fig. 4). Improvement of PGC-1α promotes the lactate metabolism and clearance. Lactate is converted to pyruvate by promoting LDH B, and MCT1 facilitates lactate uptake [45]. As shown in Table 3, the elevation of blood lactate levels was remarkably lower in the ExGPE and ExGL groups than in the ExC group. The decrease in the mRNA expression of Ldhb in the ExGPE group confirmed the decrease in lactate production by GPE supplementation (Fig. 5A). PPARδ is a nuclear receptor that regulates the fatty acid metabolism in muscles. The overexpression of PPARδ induced an oxidative fiber-type transformation and increased fatty acid oxidation in the muscle. As shown in Fig. 5B, GPE and GL supplementation in mice increased the mRNA expression of Ppard, which showed a decrease in fat mass in the ExGPE group. These results suggest that incorporating GPE may enhance fatty acid oxidation to delay the use of glucose and glycogen for the energy supply. Furthermore, GPE and GL supplementation upregulated Sod2 in the muscle (Fig. 5C). SOD2 is a major mitochondrial antioxidant enzyme that helps remove ROS. These results suggest that the anti-fatigue mechanisms of GPE are due to an increase in Sod2 mRNA expression. Fig. 7 illustrates the proposed mechanism through which GPE improves exercise performance. The beneficial effects in improving mitochondrial biogenesis may be one of the main effects in delaying muscle fatigue, enhancing muscle function, and endurance exercise performance.
Fig. 7
Summary of the effects of the G. pentaphyllum extract on the exercise endurance and energy metabolism.
GPE, G. pentaphyllum extract; p38, p38 MAP kinase; AMPK, adenosine monophosphate-activated protein kinase; PGC-1α, peroxisome proliferator-activated receptor γ coactivator 1-alpha; LDH B, lactate dehydrogenase B; LDH A, lactate dehydrogenase A; PPARδ, peroxisome proliferator-activated receptor-δ; NRF-1, nuclear respiratory factor 1; SOD2, superoxide dismutase 2; TFAM, transcription factor A mitochondrial; mtDNA, mitochondrial DNA.
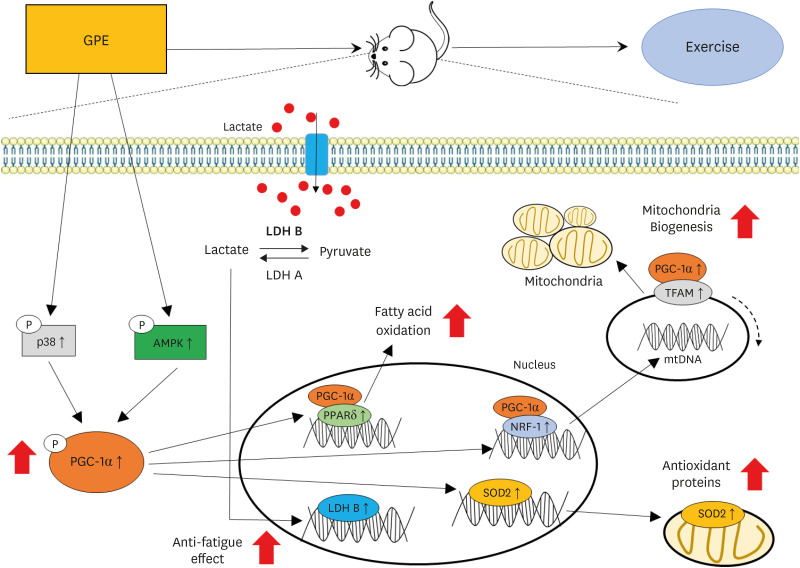
GPE supplementation upregulates PGC-1α expression in the muscle and enhances endurance exercise performance and delayed fatigue, as assessed by the supportive evidence from mRNA gene expression, to indicate the possible mechanisms for the increased serum glucose, TG, and muscle glycogen. Therefore, GPE supplementation might be used as a sports supplement candidate with improved exercise performance and anti-fatigue activity.
 XML Download
XML Download