

 PDF
PDF Citation
Citation Print
Print


Introduction
Cancer stem cells (CSCs) are a type of cell population with self-renewal and replication that have been found to be the origin of cancer (1, 2). Growing evidence indicates that CSCs are tumor-initiating cells and play a non-negligible role in cancer recurrence, metastasis, and drug resistance (1, 3, 4). However, it is unclear how CSCs are generated and maintain stemness, which make tumor treatment challenging.
Octamer-binding transcription factor 4 (OCT4), which is encoded by the Pou5f1 gene and is a member of the POU-domain transcription factor family, is connected to maintain pluripotency of embryonic stem cells (ESCs) and CSCs (5-7). Mouse Oct4 has two homologs, Oct4A and Oct4B, the former in the nucleus and the latter in the cytoplasm (8). Unlike mice, human OCT4 has three homologs, OCT4A, OCT4B and OCT4B1 (9). One study found that Oct4 expression was not detectable in adult murine organs (10). Meanwhile, some researchers also suggested that although Oct4 expression cannot be detected in testes, brain, liver, lung, kidney, and intestine, there is Oct4 expression in primordial germ cells and unfertilized oocytes, which shows that differential regulation of Oct4 expre-ssion during mouse development (11). However, a study has found that there is Oct4 mRNA expression in the murine adult ovaries and testes, especially mature and ovulating oocytes rather than resting oocytes, which may be due to Oct4 transactivate genes that are important for oocyte maturation (12). Gradually, some researchers suggest that Oct4 is necessary and sufficient to induce pluripotency of adult mouse neural stem cells (13). The expression of OCT4 in several human adult stem cells, such as breast, pancreatic, and liver stem cells, supports the hypothesis that stem cells are carcinogenic target cells (14). Additionally, OCT4 may generate resistance to radiotherapy by improving the epithelial to mesenchymal transition (EMT) process in human rectal cancer cells, and OCT4 is closely related to DNA damage when cancer cells respond to radiotherapy (15). Studies also indicate that OCT4 is strongly associated with tumor invasion and migration and can lead to poor prognosis for patients (6). Unfortunately, studies on the functions and mechanisms of OCT4 in the production of CSCs are scarce. The purpose of this review is to summarize the relationship between OCT4 and CSCs to provide possibilities for clinical treatment by exploring targets related to OCT4.
OCT4 Is a Hallmark of CSCs
The POU domain, which name derived from the Pituary-specific TF Pit1, Octamer binding TFs Oct1 and Oct2, and neural TF Unc-86, is highly conserved DNA-banding domain (16, 17). OCT4 has two DNA-binding domains, which bind to octamers of free DNA (octamer motif 5’-ATTTGCAT-3’) in a sequence-specific manner and induce chromatin opening and regulate gene expression (17, 18). The two DNA-binding domains consist of a POU-specific domain (POUS) and a POU homeodomain (POUHD) (19). Compared with other stem genes, OCT4 has universal expression in CSCs of hepatocellular carcinoma, breast cancer, prostate cancer, melanoma, osteosarcoma, bladder cancer, ovarian cancer, and lung cancer (7, 20, 21). Oct4 is overexpressed in pluripotent embryonic cells and silenced after cell differentiation during mouse embryonic development (9). It seems that OCT4 can be a pluripotency and germ cell marker and be used to distinguish from non-CSCs, and participate in determining the biological function of CSCs.
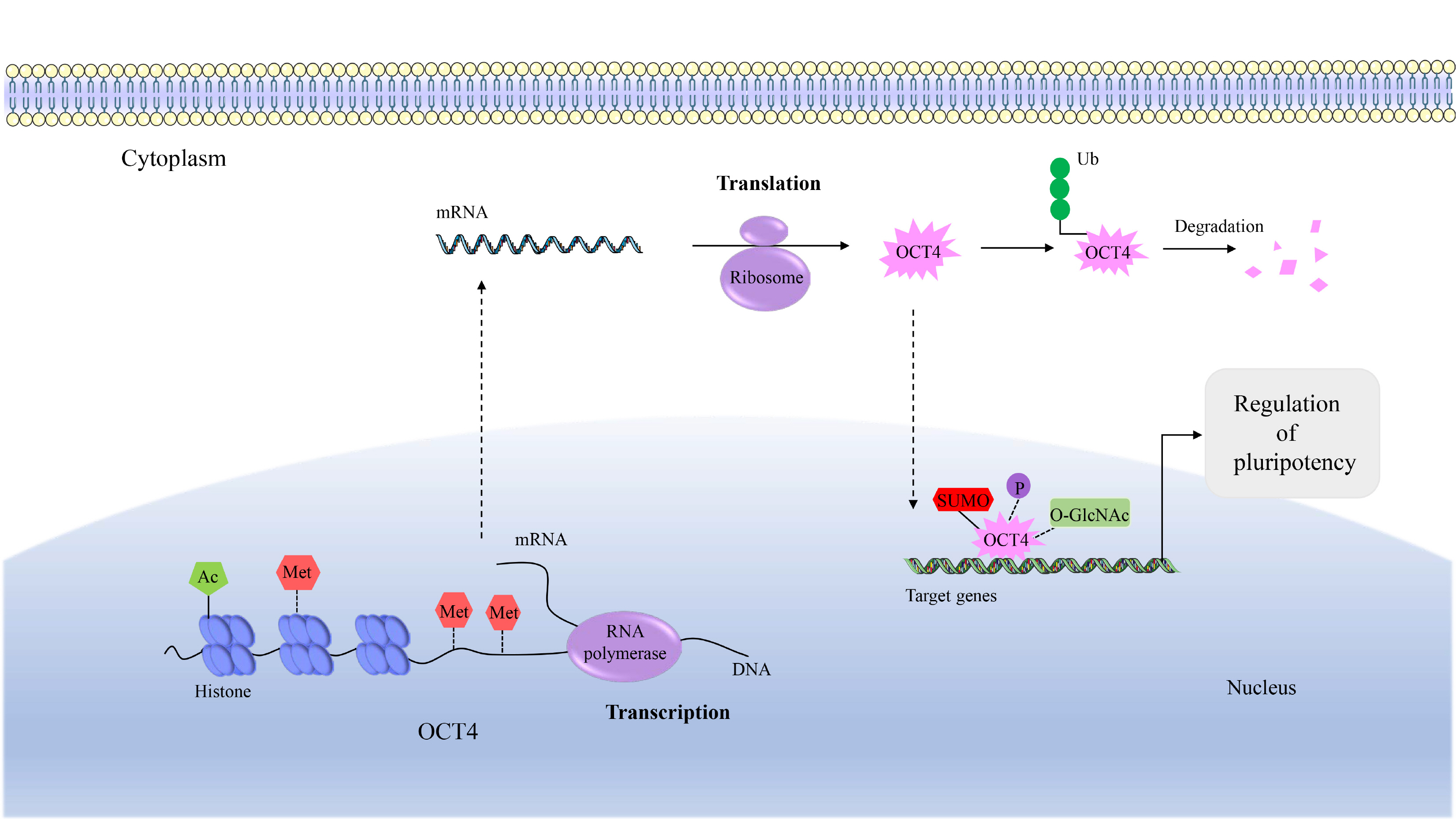
Regulation of OCT4 Expression by Epigenetic Modification
Epigenetic modification is a reversible and heritable change in gene function when the DNA sequence has not changed, which can participate in important biological processes by regulating gene expression (22). OCT4 silencing plays important role in differentiation, cell engineering, and tumors (23). In contrast, OCT4 expression is related to stem cell characteristics and increases spheroid formation capacity (24).
Methylation
Studies have shown that OCT4 is regulated by DNA methylation of CpG of the promoter and exon in the human trophoblast cells and ESCs (25, 26). Hypermethy-lation of the Oct4 promoter and enhancer regions results in structural changes in chromatin and inhibits Oct4 expression in mouse trophoblast stem cells (27). However, the epigenetic regulation of the histones in the OCT4 promoter region cannot be ignored. The OCT4 promoter region includes the CCCTC-binding factor (CTCF) binding site. As it happens, Brother of the Regulator of Imprinted Sites (BORIS) as a CTCF paralog can bind to the OCT4 promoter, and it promotes OCT4 expression and promotes stemness of human hepatocellular carcinoma by up-regulating H3K4me2 and down-regulating the level of H3K27me3 (28).
Acetylation
Researchers indicated that acetylation of OCT4 and SOX2 can attenuate transcriptional activity by impairing OCT4/SOX2 heterodimer formation (29). The expression of OCT4 is positively correlated with Ca2+/calmodulin-dependent protein kinase IIγ (CaMKIIγ), which promotes the acetylation of the histones of OCT4 by activating Akt, thereby maintaining the stem cell capacity and tumorigenicity of human lung cancer cells (30). H3K56 acetylation is highly conserved in organisms, which can interact with Oct4 and promote the pluripotency of mouse ESCs (mESCs) (31). A significant increase in the acetylation level of H3K9 in the OCT4 promoter region was found in spherical cultures formed from human mesenchymal stem cells (32), which shows that the post-translational modification of OCT4’s histones plays an important role in stem maintenance.
Phosphorylation
Phosphorylation that occurs in the POUHD region of OCT4 in human ESCs (hESCs) inhibits its activity by blocking sequence-specific DNA binding (19). Studies have also found that Akt-mediated OCT4 phosphorylation can regulate stemness by changing the interaction with SOX2 (29). And increasing evidence suggests that phosphorylation at threonine 343 of Oct4 is also critical for maintaining mESC pluripotency (33). In addition, different genetic modifications also interact with each other. The phosphorylation of human OCT4 at serine 111 promotes its ubiquitination, which affects its activity and cell localization (34).
Ubiquitination, SUMOylation, and glycosylation
Experiments show that E3 ubiquitin ligase WWP2 promotes the ubiquitination and degradation of OCT4 in hESCs. The same result is also observed in differentiated mouse embryonic carcinoma cells. Furthermore, ITCH, another ligase, regulates mESC Oct4 transcription and degradation after ubiquitination (9, 34, 35). Carboxy terminus of HSP70-interacting protein (CHIP), which is an E3 ubiquitin ligase, can ubiquitinate OCT4 at lysine 284, which can reduce OCT4 stability and subsequently inhibit human breast CSC production (36). Oct4 SUMOylation in mouse embryonic carcinoma cells results in enhanced protein stability, transactivation function and DNA binding (7, 29). Studies have found that OCT4 expression increases in cells of rectal cancer, neuroblastoma, and melanoma after drug treatment. The difference is that testicular germ cell tumors treated with cisplatin appear to reduce OCT4 expression through SUMOylation, but specific mechanism is still unclear (37). There is evidence that the protein activity of human OCT4 is altered by monosaccharide O-linked β-N-acetylglucosamine (O-GlcN Ac) (9). O-GlcNAc at the threonine 228 of Oct4 boosts the transcription of Oct4 and thus induces a variety of pluripotency genes, which is essential for the reprogramming of mESCs (38).
In summary, the same genetic modification may regulate the structure and function of OCT4 through different pathways due to the role of different enzymes. Various genetic modifications participate in the regulation of OCT4 activity, stability and cell localization through interaction in various forms (Fig. 1), all of which have an effect on the role of OCT4 and provide new perspectives and targets for cancer treatment.
Non-Coding RNA Related to OCT4
There are many types of non-coding RNA (ncRNA), such as long non-coding RNA (lncRNA), small interfering RNA (siRNA) and micro RNA (miRNA, miR) (39). Non-coding RNA is involved in cell growth, differentia-tion, apoptosis, invasion, and other important biological processes, and even some of them have proven to be crucial in the development of cancer (40, 41). OCT4, as a representative CSC marker, is often used to judge the role of ncRNA in CSCs, such as miR-30b, lncRNA HOXA11-AS, and lncRNA MEG3 (42-44). In addition, more and more ncRNAs have been shown to be related to the regulation of CSCs by interacting with OCT4.
Long non-coding RNA (lncRNA)
OCT4 binds to the lncRNA NETA1.1 promoter in human bladder cancer-resistant cells treated with cisplatin, making it highly expressed to maintain the invasion and growth of bladder cancer cells (45). LncRNA CCAT2 overexpressed in human breast CSCs maintains the aggressiveness of CSCs by up-regulating the OCT4 pseudogene (OCT4-PG1) (46). The ability of MALAT1 to maintain the stemness of human colon CSCs may be achieved by targeted inhibition of miR-20b-5p and reducing the binding of miR-20b-5p to OCT4 mRNA (Pou5f1) (47). OCT4 promotes the invasion and proliferation of cancer cells by regulating the level of lncRNA AK055347 in human osteosarcoma cells (48). The activation of lncRNA AK028326 was also found to be directly regulated by Oct4 in mESCs (9). Lnc-CRCMSL, as an anti-metastatic gene, prevents human colorectal cancer cells from reprogramming by inhibiting high mobility group box 2 (HMGB2) from metastasizing to the nucleus and inhibiting the interaction between HMGB2 and OCT4 (49). H19 can directly regulate the expression of OCT4 in human prostate cells. In turn, in F9 embryonic carcinoma cells, Oct4 and Sox2 can also positively regulate H19 by weakening the methylation level of the imprinted control region and promoter (50, 51). And research has also found that lncRNA ROR can form a regulatory feedback loop with OCT4, SOX2, and NANOG in hESCs (52, 53).
Micro RNA (miRNA, miR)
Further evidence suggests that the involvement of miRNA in maintaining pluripotency is due to the presence of OCT4 binding sites on miRNA promoters (54). It is worth noting that OCT4, SOX2 and NANOG can bind to the promoter of miR-302 and promote its expression in hESCs (55). The miR-302 can also indirectly positively regulate OCT4 activity to promote reprogramming efficiencies from human adipose-derived stem cells into induced pluripotent stem cells through targeted inhibition of Nuclear receptor subfamily 2, group F, member 2 (NR2F2) that suppresses OCT4 promoter activity (56). MiR-145 directly targets the 3’UTR of OCT4, which inhibits the transcription of OCT4 and promotes the differentiation and inhibits the proliferation of human endometrial adenocarcinoma cells (57), and the same result was also found in glioblastoma (58). Coincidentally, OCT4 also binds to the promoter of miR-145 to achieve functional inhibition of miR-145, which seems to indicate that a double-negative feedback loop can be formed between miR-145 and OCT4 in hESCs (9, 59). The lncRNA linc-DYNC2H1-4, which present in the cytoplasm of gemcitabine-resistant pancreatic cancer cells, competes with miR-145 and subsequently eliminates the targeted inhibi-tion of OCT4 by miR-145 to promotes CSC phenotypes (60). Studies in endometrial cancer and hepatocellular carcinoma have still found that the targeted inhibition of OCT4 by miR-145 can be reversed by OCT4 pseudogene 5 (OCT4-pg5) due to their similar binding sites in OCT4 3’UTR (61, 62). In addition, a decrease of lincRNA ROR in prostate CSCs can increase the effective concentration of miR-145 and inhibit CSC proliferation (50, 63). In short, many ncRNAs may use miR-145 as an intermediate to complete the connection with OCT4. MiR-302/367 cluster regulated by OCT4 plays a role in maintaining pluripotency of stem cells (54). OCT4 directly regulates miR-1246, and they collectively activate the Wnt/β-catenin signaling pathway, which can promote self-renewal, tumorigenicity and drug resistance of liver CSCs (64).
Small interfering RNA (siRNA)
Using siRNA to target OCT4 may become a means to eradicate CSCs, which can achieve the purpose by inducing CSCs to age and apoptosis (65). The discovery of siOCT4 in head and neck squamous cell carcinoma CSCs inhibits EMT and resistance by targeting the increased OCT4 in CSCs, which proves a new possibility for eradicating cancer cells and reducing metastasis and recurrence (66). The same siRNA-mediated study of OCT4 targeted silencing has also been found in pancreatic cancer, showing inhibition of pancreatic cancer cell proliferation and induction of apoptosis (67). The use of OCT4 siRNA in breast CSCs reduces drug resistance to paclitaxel and tumor initiating ability (68). However, the efficiency of siRNA delivery is limited and needs to be improved. But the targeted therapy of OCT4 still provides us with new ideas for CSC treatment, which may need us to study more comprehensive technologies.
In fact, many ncRNAs can affect the expression of OCT4 when regulating CSCs, but the ncRNAs that actually interact with OCT4 are somewhat scarce. And the interaction between ncRNA and OCT4 is extremely complicated (Table 1). The ncRNAs can directly bind to OCT4, or form a regulatory network with OCT4 through an “intermediate”, which provides multiple therapeutic targets for eradicating CSCs.
OCT4 Participates in the Formation of Complexes
Protein complexes are complexes formed by two or more functionally related proteins through disulfide bonds or other interactions, which have a huge influence in the occurrence and progression of cancer, such as cell localization, gene transcription, DNA damage repair, cell cycle, cell differentiation, and other biological processes (69, 70). OCT4 forms protein complexes with different partners (SOX2, NANOG, KLF4 or other proteins) to participate in the regulation of proliferation and self-renew of CSCs (29, 71).
OCT4/SOX2 protein complex
Of these, the OCT4/SOX2 protein complex is common (72). The Oct4/Sox2 complex single-molecule imaging model indicates that Sox2 may first bind to the chromosome, which provides a target for subsequent Oct4 binding, and that Oct4 binding can stabilize the Oct4/Sox2 structure in mESCs (73). The OCT4/SOX2 complex induces transcription in the nucleus and leads to the concept that binding partners can stimulate nuclear localization (8). And they bind to specific target genes to induce their expression and jointly maintain CSC-like characteristics. However, whether the OCT4/SOX2 complex works may be related to post-translational modification and the specific mechanism is not clear (5, 7, 9). Subsequently, it was discovered that OCT4/Lys-156 has different post-translational modifications in human pluripotent stem cells and differentiated cells, which may affect the stability of the OCT4/SOX2 complex and regulate the EMT phenotype (73). In the structure of the OCT4/SOX2 complex, the salt bridge formed between OCT4/Lys-151 and SOX2/Asp-107 is relatively obvious, so post-translational modification or mutation of key residues may destroy the salt bridge structure and damage the stability of the protein complex, which seems to reduce the maintenance of stem cell-like characteristics by mechanically damaging the complex structure (73). The OCT4/SOX2 complex has also been shown to be involved in DNA repair, cell cycle, and apoptosis (74). Zfp206 as a transcrip-tion factor involved in maintaining pluripotent stem cells, has been shown to interact with the Oct4/Sox2 complex in mESCs and is an important part of the complex (9, 75). In head and neck squamous cell carcinoma, OCT4/SOX2/NANOG may form complexes or regulatory networks to prevent differentiation and lead to poor prognosis and chemoresistance (71).
Other protein complexes
β-catenin forms complexes with Oct4 and Klf4 and participates in regulating stem cell-like characteristics in mESCs (55). The sal-like 4 (SALL4) as a transcription factor maintains the proliferation, chemoresistance and self-renewal ability of CSCs and can form a complex or regulatory network with OCT4 to maintain CSC-like characteristics (76-78). Ku80 encoded by the XRCC5 gene is related to the repair of double-stranded DNA breaks and can interact with SALL4, thereby interfering with the stability of the SALL4/OCT4 complex and destroying the self-renewal ability of liver CSCs (76). The scaffolding protein caveolin-1 (Cav-1), which has a tumor suppressive effect and is low-expressed in the tumorspheres of breast cancer, can form a complex with OCT4 and mediate the degradation of OCT4 through ubiquitin-proteasome (79, 80). However, NO can damage the Cav-1/OCT4 complex by promoting phosphorylation on Cav-1 tyrosine 14 and boost the stability and biological function of OCT4 and lung CSC-like phenotype (79, 81). Under hypoxic or glucose-restricted conditions, nuclear PKM2, as an isozyme of pyruvate kinase, can bind to OCT4 and collectively regulate the transcription of downstream stemness-related genes, thereby increasing cancer invasion and metastasis (82, 83). The BAF (BRG1-associated factor) complex interacts with the Oct4 to form the Oct4/BAF complex, which regulates epigenetic modifications of mESC differentia-tion (84). Functional proteins can also combine with each other on the OCT4 gene to form a complex. For example, the transcription factor ZIC2 recruits nuclear remodeling factor (NURF) to form a complex in the OCT4 promoter region, activates OCT4 transcription, and promotes the self-renewal and differentiation potential of liver CSCs (85).
With the interaction between W118 residue of Ash2I and Oct4, Ash2I can further recruit other transcription factors Sox2 and Nanog to form the Ash2I/OSN complex in mESCs and activate downstream stem genes to jointly regulate the pluripotent network by epigenetic modifications (86). Increased ABCG2 (ATPase binding cassette transporter protein) in glioblastoma stem cells may interact with OCT4 to promote drug resistance and CSC survival (87, 88). The complexes formed by OCT4 may induce other pluripotency-related transcription factors to regulate the characteristics of CSCs. For example, the Oct4/Sox2/Klf4 complex binds to the Nanog promoter to induce its transcriptional activity and achieve the function of regulating stem cell-like characteristic in mESCs (89). Actually, some researchers have suggested that OCT4 is sufficient for pluripotential reprogramming of human neural stem cells (26).
OCT4 can independently perform stem-related regulation or form a complex with stem cell transcription factors or other functional proteins to regulate CSC proliferation, self-renewal, and invasion (Table 2). Therefore, how OCT4 interacts with binding proteins will become our research direction, which may provide new methods for our CSC treatment by interfering with complex formation or blocking regulatory networks.
Signaling Pathways Related to OCT4
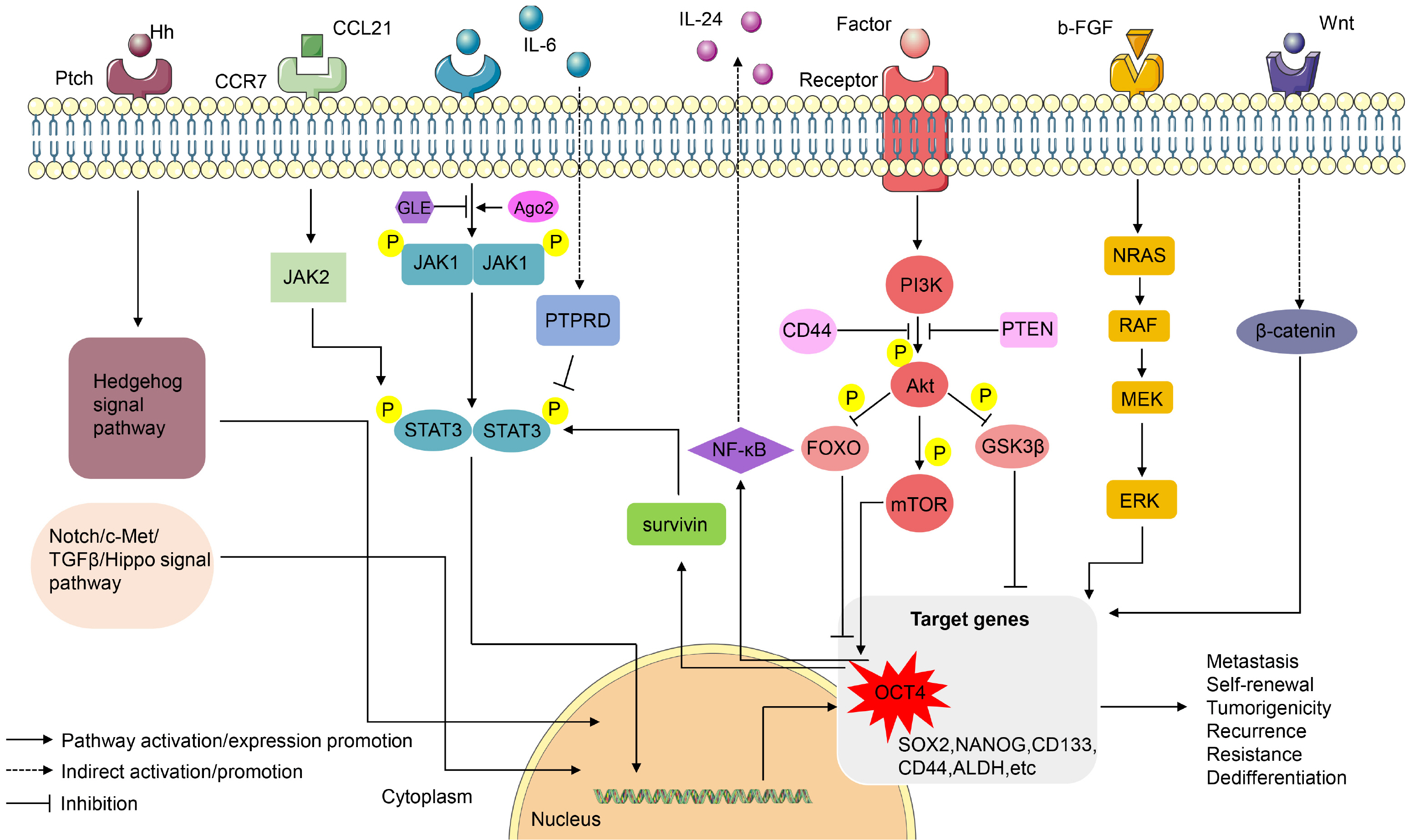
In addition to being regulated by pluripotency-related transcription factors (such as OCT4, SOX2, NANOG, KLF4, MYC) and extracellular factors (such as hypoxia and extracellular matrix), CSCs are also interfered by various signaling pathways (such as Wnt, Notch, PI3K/Akt, JAK/STAT, Hedgehog) (90). Studies have found that OCT4 is also involved in the complex regulation of CSCs as an intermediate station or terminal target in the signaling pathway.
Hedgehog signaling pathway
Hedgehog signaling pathway stimulated by the binding of Hh ligand and transmembrane protein receptor PTCH can regulate CSC metastasis and self-renewal ability by up-regulating the expression of downstream target gene OCT4 (52, 90). The lncRNA-Hh regulated by the gene TWIST, induces the expression of OCT4 by promoting the Hedeghog pathway and maintains the tumorigenicity and self-renewal ability of breast CSCs (52).
Signal transducer and activator of transcription pathway
Increasing evidence suggests OCT4 is functionally dependent on signal transducer and activator of transcription 3 (STAT3) and the expression of OCT4 is positively related to the activity and expression of STAT3 in breast CSCs, cervical CSCs and liver CSCs (91-95). Inter-leukin 6 (IL-6) activates the Janus kinase1 (JAK1)/STAT3 signaling pathway and downstream OCT4 expression to complete the transformation of human breast cancer cells into breast CSCs (96, 97). The leukemia inhibitory factor (LIF) as a member of the IL-6 family can also activate this pathway (97). And IL-6 can also upregulate OCT4 through JAK2/STAT3 signaling to maintain the recu-rrence and drug resistance of liver CSCs (98). Interestingly, IL-6 also transiently upregulated protein tyrosine phosphatase receptor-type δ (PTPRD), which in turn dephosphorylates STAT3 and inhibits IL-6/STAT3 signal transduction and OCT4 expression in human breast cancer cells (99). At the same time, it was found that OCT4 can also activate the JAK1/STAT6 signaling pathway in ovarian CSCs and promote tumorigenesis (90). Moreover, OCT4 induces the production of IL-24 through the STAT3 and NF-κB signaling pathways, which can confer radiotherapy resistance to breast cancer cells by inhibiting radiation-induced senescence (65). The recognition and combination of ligand chemokine ligand 21 (CCL21) and C-C chemokine receptor 7 (CCR7) increase the expression of pluripotency-related transcription factors and stem cell markers (such as OCT4, CD133, CD44) by activating the JAK2/STAT3 signaling pathway, thereby enhancing the invasion, migration, and tumorsphere formation of oral squamous cell carcinoma (100). In addition, the STAT3 or STAT5A plays an important role in glioblastoma, which can be verified by the expression of OCT4 (101). The study further found that JAK/STAT3 activity is essential for DNA demethylation of Oct4 promoter during mouse somatic cell reprogramming (102, 103). Ganoderma lucidum extract (GLE) with anticancer activity inhibits the STAT3 signaling pathway thereby reducing the expression of phosphorylated and total STAT3 and OCT4 of breast CSCs (91). Ovatodiolide reduces the expression of genes, such as OCT4, and the formation of tumorspheres by inhibiting the JAK2/STAT3 signaling pathway, and cooperates with cisplatin to complete the treatment of oral cancer (104). Survivin is an apoptosis suppressor protein and is associated with chemoradiation resistance and metastasis of tumor cells (105). In hepatocellular adenocarcinoma, OCT4 was found to regulate the migration and invasion of cancer cells through the Survivin/STAT3 signaling pathway (106).
PI3K/Akt pathway
The PI3K/Akt pathway, which is related to cell growth and apoptosis, phosphorylates the substrate FOXO transcription factor to inactivate it and thereby reducing the expression of OCT4, because FOXO can directly bind to the OCT4 promoter to regulate OCT4 transcription, which indicates that targeting FOXO factors can reduce the generation of CSCs and the use of PI3K inhibitors clinically has potential risks (107-109). Likewise, phosphorylated Akt can phosphorylate OCT4, which increases tumorigenicity and self-renewal ability of CSCs (110). However, the researchers demonstrate that the Akt pathway is activated when mouse embryonic carcinoma cells begin to differentiate, and Akt phosphorylates Oct4 at serine 228 to accelerate its degradation (111).
Wnt/β-catenin signaling pathway
The Wnt/β-catenin signaling pathway plays a huge role in maintaining the stem cell-like characteristics of breast CSCs and ESCs (112, 113). Diallyl Trisulfide can suppress Wnt/β-catenin signaling pathway and OCT4 expression to inhibit breast CSCs (112). Exogenous intake of bisphenol A (BPA) and polychlorinated biphenyls (PCBs) can increase the expression of stem cell markers, such as OCT4, and drug resistance of human ovarian cancer cells by activating Wnt/β-catenin pathway (114). OCT4 can also bind to enhancers of target genes activated by the Wnt/β-catenin signaling pathway, so the deletion of OCT4 has a direct influence on the Wnt/β-catenin signaling pathway (115). A contradictory result suggests that Wnt/β-catenin plays a role in hESC differentiation rather than self-renewal and that OCT4 may inhibit this pathway (116).
Other signaling pathways
Studies have found that fine particulate matter (PM2.5) exposed to human lung cancer cells promotes the expression of OCT4 by activating the Notch signaling pathway, thereby promoting the occurrence of lung CSCs (117). Similarly, the relationship between the Notch pathway and OCT4 has also been confirmed in pancreatic CSCs (118). Evidence suggests that the c-Met signaling pathway promotes the self-renewal and metastasis of glioblastoma stem cells by up-regulating the expression of OCT4 and c-MYC (119). TGF-β RI is highly expressed in tissues with highly metastatic endometriosis, and it was found that the completion of TGF-β signal transduction requires the addition of OCT4 (120). Moreover, the knockout of OCT4 in liver cancer cells significantly reduces the expression of genes related to the TGF-β pathway (ELF, Smad3, Smad4), which indicates that OCT4 plays an important role in CSCs by improving the TGF-β pathway (121). The Hippo signaling pathway was found to be inhibited during the progression from colorectal adenoma to colorectal cancer, and OCT4 as a target gene of the Hippo pathway was upregulated in the case of overexpression of downstream cascade kinases, which may be related to the progression and metastasis of colorectal cancer (122). The pluripotency mediator b-FGF upregulates the expression of OCT4 and maintains the undifferentiated state of human induced pluripotent stem cells through the mitogen-activated protein kinase (MAPK) signaling pathway (NRAS-RAF-MEK-ERK) (123).
Collaboration between signaling pathways
The various signaling pathways are not independent, and they may be synergistic in regulating OCT4 expression. Argonaute 2/OCT4/methyl-CpG-binding protein 6 (Ago2/OCT4/MBD6) signal transduction pathway was found to regulate stemness-related genes and human adipose tissue-derived stem cell self-renewal (124). In addition, Ago2 can also regulate human umbilical cord blood-derived me-senchymal stem cell self-renewal through the expression of OCT4 and activation of Wnt/β-catenin and JAK2/STAT3 signaling pathways (125). IL-17B related to cancer progression enhances the expression of OCT4, SOX2 and other transcription factors and activates the NF-κΒ, STAT3 and β-catenin pathways to promote the progre-ssion of gastric cancer (126). IL-23, which is positively correlated with OCT4 expression, promotes the self-renewal ability and tumorigenicity potential of ovarian CSCs through NF-κΒ and STAT3 signaling pathways (127). The PI3K/Akt2/mTOR signaling pathway and MAPK signaling pathway promote the cisplatin and radiation resistance of neuroblastoma cells by regulating the expression of OCT4, SOX2, CD133 and ALDH (128). In addition to the PTEN/PI3K/AKT/β-catenin axis, miR-429, which is hypomethylation and highly expressed in liver CSCs, also regulates the generation, invasion and metastasis of liver cancer through the Rb binding protein 4/E2F transcription factor 1/OCT4 (RBBP4/E2F1/OCT4) axis (129, 130). Phosphorylation and inactivation of GSK3β by Wnt, PI3K, Akt, and MAPK pathway reduce the expression of OCT4, but in head and neck cancer, CD44 can inhibit Akt phosphorylation and thus inhibit GSK3β inactivation and maintain the self-renewal ability of CSCs (131).
In fact, OCT4-related signaling pathways are complex in maintaining the stem characteristics of CSCs (Fig. 2). There may be conflicting research results on the same signal pathway, which requires us to further determine whether there is a difference in cancer species specificity or cellular time limit. The discovery of signal pathways related to OCT4 provides many targets for radical treatment of CSCs, which may well solve the problems of cancer recurrence, metastasis and drug resistance.
Cancer Treatment Effects with OCT4 Targeted
The role of OCT4 in pluripotency maintenance makes OCT4 a new cancer treatment target. Increased experiments have found that siRNA targeting OCT4 induces apoptosis, inhibits proliferation, EMT, and drug resistance in pancreatic cancer cells and head and neck squamous cell carcinoma CSCs, breast CSCs (66-68). Similar results have been found in lung cancer, ovarian cancer, liver cancer, and glioma (37, 106). However, an experiment suggested that OCT4 knockout in the MCF-7 human breast cancer cell line induced cell invasion, migration, and EMT, which is because MCF-7 cells already have high expression of OCT4 (37, 132). Moreover, due to the intra- and extracellular degradation of enzymes, the delivery efficiency of siRNA is limited, which still needs to be solved (66). The epigenetic regulator JMJD3 inhibits OCT4 expression in human breast cancer cells in an independent manner with demethylase activity, and paricalcitol, a vitamin D analog, inhibits OCT4 expression and stem cell-like characteristics of human breast cancer cells after promoting JMJD3 expression (133). Recently, three different Oct4 epitopes were chemically synthesized, among which Oct4-3 and carrier protein KLH induced a strong tumor-specific adaptive immune response in combination with toll-like receptor 9 agonist, thereby inhibiting mouse testis embryonic carcinoma growth and promoting long-term survival. Importantly, the mice were well tolerated with the Oct4-3 vaccine and no obvious adverse events were observed (134). Additionally, targeting upstream activators of OCT4 is also a new method. For instance, the use of Notch pathway inhibitor L685,458 can reduce the expression of OCT4 and reverse stem cell-like phenotype and resistance to paclitaxel of breast cancer cells (19, 135). Using siRNA to target OCT4B1 induces apoptosis or G2/M arrest in human brain cancer cells, suggesting that OCT4B1 may also be a potential therapeutic target in brain cancer (136).
Conclusions
CSCs are a group of cells with the ability to self-renew and differentiate among cancer cells. Their discovery provides the possibility to solve tumor recurrence, metastasis and resistance. OCT4 as a stem transcription factor is widely expressed in ESCs and CSCs, and research has found that OCT4 may participate in the regulation of CSCs through various forms, but the specific mechanism may not be clear. This review mainly proposes the specific role of OCT4 in CSCs from the epigenetic modification of OCT4 and complexes, non-coding RNA and signaling pathways related to OCT4, which may provide multiple targets for the treatment of CSCs to achieve the purpose of extending patient life.
 XML Download
XML Download