

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Obesity is an increasingly worrisome public health issue and is associated with several leading causes of morbidity and mortality, such as cardiac diseases, diabetes, and spleen and liver diseases [1]. Obesity is one of the most noteworthy conditions that can increase the risk of a poor prognosis in many immune-related conditions [2]. The spleen is an immune system organ that has a crucial role in the primary host defense immune response against bacterial invasion [3]. Despite being a critical organ, the relationship between a high fat/cholesterol diet (HFCD) and splenic inflammation risk factors in mice has not been fully described.
Activation of nuclear factor kappa B (NF-κB) leads to its translocation into the nucleus, where it binds to κB sites. It stimulates the production of pro-inflammatory cytokines such as interleukin 6 (IL-6) and induces the expression of other related pro-inflammatory genes, including cyclooxygenase 2 (COX-2) and inducible nitric oxide synthase (iNOS) [4]. Growing experimental evidence from in vitro and in vivo studies suggests that NF-κB is an inflammatory mediator [5]. Toll-like receptors (TLRs) are essential in innate immune processes and metabolic disorders including obesity [6]. The TLR4 signaling pathway is central to the inflammatory response induced by obesity [7]. Myeloid differentiation primary response 88 (MyD88), a key intracellular mediator of inflammatory signaling, is an essential signaling adapter for TLRs and is associated with metabolic disorders linked to obesity [78]. Therefore, the TLR-MyD88 pathway is crucial in HFCD-induced splenic inflammation.
Recently, many researchers have focused on natural dietary supplements considered beneficial for health. In particular, evidence of the anti-obesity effects of such supplements is accumulating from both in vivo and ex vivo studies [910]. Natural compounds containing phytochemicals such as epigallocatechin gallate [11], curcumin [12], and quercetin [13] have been shown to exhibit anti-adipogenic and anti-inflammatory effects.
Several studies have shown that consumption of cruciferous vegetables such as cauliflower, cabbage, and broccoli can lead to chemoprevention in various rodent models [14]. Glucosinolate is a sulfur-containing compound present in cruciferous vegetables. Phenethyl isothiocyanate (PEITC) is an active ingredient of cruciferous vegetables and has been extensively studied for its various anti-cancer and tumor cell inhibitory effects [15]. In addition, 3,3′-diindolylmethane (DIM) is present in cruciferous vegetables, especially brassica species, such as cabbage, Chinese cabbage, and broccoli [16]. Several studies have indicated that DIM has anti-cancer and anti-oxidant properties [1718]. In our previous study, we observed that a HFCD can induce obesity and inflammation of liver tissue in C57BL/6 mice [19]. However, we have not determined whether a HFCD increases inflammation by examining serum and spleen tissue samples. Also, the effects of PEITC and DIM on inflammation in immune processes in spleen tissue have not been elucidated. Therefore, this study aimed to investigate the anti-inflammatory effects of PEITC and DIM on the spleen of C57BL/6 mice being fed a HFCD.
MATERIALS AND METHODS
Animals and diet
Four-week-old C57BL/6 mice were purchased from Orient Bio Co. (Seongnam, Korea). After acclimatizing for one week, the mice were randomly divided into six groups (n = 10). PEITC and DIM were purchased from Sigma-Aldrich (St. Louis, MO, USA). The groups were fed a normal AIN-93G diet (CON group), a high fat diet (60% calories from fat) with 1% cholesterol (HFCD group), a HFCD with 30 mg/kg/day PEITC (HFCD+P30 group), a HFCD with 75 mg/kg/day PEITC (HFCD+P75 group), a HFCD with 1.5 mg/kg/day DIM (HFCD+D1.5 group), or a HFCD with 7.5 mg/kg/day DIM (HFCD+D7.5 group). The mice were supplied their diet and water ad libitum and were housed in a stable temperature-controlled environment with a 12 h light/dark cycle. Mouse body weight was monitored every three days. After sacrificing the mice, spleen tissue was immediately excised, weighed, and stored at −80°C. The experimental protocol was approved by the Animal Care and Use Committee of Chonnam National University (CNU IACUC-YB-2019-01).
Preparation of nuclear fractions
The cytoplasmic lysate was extracted in cytosolic lysis buffer with 10% NP-40. The lysis buffer for the cytosolic lysate was composed of 10 mM HEPES, 10 mM KCl, 0.1 mM EDTA, 0.1 mM EGTA, 1 mM dithiothreitol, and 0.5 mM phenylmethylsulfonyl fluoride (PMSF). The cytoplasmic lysates were centrifuged, and the supernatant was collected as the cytoplasmic fraction. The remaining pellet was then re-suspended in nuclear extraction buffer with 10% NP-40. The extraction buffer for the nuclear lysate was composed of 20 mM HEPES, 0.4 mM NaCl, 1 mM EDTA, 1 mM EGTA, 1 mM dithiothreitol, and 1 mM PMSF. The nuclear lysates were collected, cleared by centrifugation at 16,343 × g for 1 min at 4°C, and the supernatant was aliquoted and stored at −80°C.
Western blotting
Protein samples (40 μg) were mixed with sample buffer (100 mM Tris pH 6.8, 2% sodium dodecyl sulfate, 1% 2-mercaptoethanol, 2% glycerol, and 0.01% bromophenol blue). Total proteins were loaded onto 10% polyacrylamide gel and electrophoresis was performed using the Mini-Protean Tetra Cell system (Bio-Rad, Hercules, CA, USA). The resolved proteins were transferred to nitrocellulose membranes (Merck Millipore Ltd., Tullagreen, Carrigtwohill, County Cork, Ireland), which were then incubated for 2 h in a blocking buffer containing 5% nonfat milk. The blots were incubated with NF-κB p65 for 2 h. After incubation with a primary antibody, the blots were washed twice and incubated with the appropriate secondary antibody for 2 h. The blots were washed three times and treated with Western Blotting Luminol Reagent (Santa Cruz Biotechnology, Dallas, TX, USA), followed by autoradiography using the ChemiDoc XRS+ System (Bio-Rad). Blots were confirmed via stripping and then reprobing with Histone H3. The concentrations of antibodies used in this study were as follows: 1:500 for NF-κB p65 and 1:1000 for Histone H3. Antibodies for NF-κB p65 were purchased from Santa Cruz Biotechnology, and that for Histone H3 was purchased from Cell Signaling Inc. (Beverly, MA, USA).
Quantitative polymerase chain reaction (qPCR)
Total RNA was prepared using TRIzol reagent (Ambion, Life Technologies, Carlsbad, CA, USA) according to the manufacturer's instructions. The cDNA was synthesized from 1 μg of total RNA using the Omniscript RT kit (QIAGEN, Hilden, Germany). SYBR green-based qPCR was performed with the CFX96 Touch Real-Time PCR Detection System and the iQ SYBR Green Supermix (Bio-Rad), according to the manufacturer's instructions. Data were analyzed using the 2−ΔΔCT method using β-actin mRNA as the normalization control gene. The qPCR reaction consisted of three steps: the first step (3 min at 95°C) activated the polymerase; the second step comprised 40 cycles of denaturation (10 s at 95°C) followed by annealing (30 s at 57°C); the third step was performed to generate temperature dissociation curves of the products by incubation for 1 min at 95°C, 5 s at 65°C, and 5 s at 95°C. The primers, designed with an online program (Bioneer, Daejeon, Korea), were as follows (Forward, Fw; Reverse, Rv): mouse interleukin 6 (IL-6) Fw 5ʹ-TACCACTTCACAAGTCGGAGGC-3ʹ Rv 5ʹ-CTGCAAGTGCATCATCGTTGTTC-3ʹ, mouse COX-2 Fw 5ʹ-TGAGTACCGCAAACGCTTCTC-3ʹ Rv 5ʹ-TGGACGAGGTTTTTCCACCAG-3ʹ, mouse NF-κB p65 Fw 5ʹ-ACCACTGCTCAGGTCCACTGTC-3ʹ Rv 5ʹ-GCTGTCACTATCCCGGAGTTCA-3ʹ, mouse TLR2 Fw 5ʹ-TCCCTTGACATCAGCAGGAACACT-3ʹ Rv 5ʹ-GCAGCCGAGGCAAGAACAAAGAAA-3ʹ, mouse TLR4 Fw 5ʹ-GGCAGCAGGTGGAATTGTAT-3ʹ Rv 5ʹ-AGGCCCCAGAGTTTTGTTCT-3ʹ, mouse MyD88 Fw 5ʹ-ACTGGCCTGAGCAACTAGGA-3ʹ Rv 5ʹ-CGTGCCACTACCTGTAGCAA-3ʹ, mouse β-actin Fw 5ʹ-ACTGCCGCATCCTCTTCCTC-3ʹ Rv 5ʹ-CTCCTGCTTGCTGATCCACATC-3ʹ.
Enzyme-linked immunosorbent assay (ELISA)
Immediately after sacrifice, mouse serum was collected and stored at −80°C until assay. Inflammatory factor serum IL-6 was measured using an ELISA kit (Raybio, Norcross, GA, USA). All procedures were performed according to the manufacturer's protocol.
Statistical analysis
Each experiment was performed at least three times. Data are expressed as mean ± SD values. The significance of differences between groups was determined by applying a one-way analysis of variance (SPSS version 25.0 software, SPSS Institute, Chicago, IL, USA), while Duncan's multiple range test was used to compare significant differences between groups. Statistical significance was present at P-values < 0.05.
RESULTS
Changes in spleen weights in HFCD-fed mice
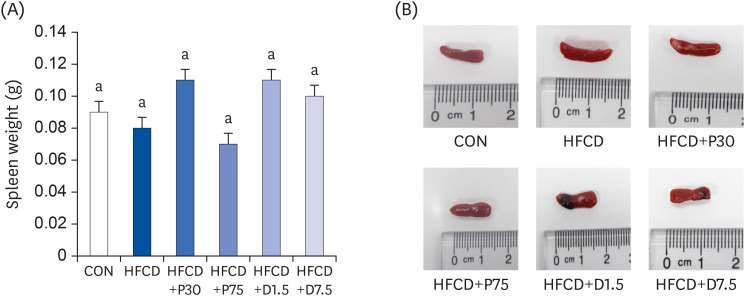
The changes in mouse spleen weights during the entire 13-week feeding period are shown in Fig. 1A and B. Spleen weights of the CON and HFCD groups were 0.09 ± 0.02 g and 0.08 ± 0 g, respectively. Spleen weights of the HFCD+P30 and HFCD+P75 groups were 0.11 ± 0.02 g and 0.07 ± 0.01 g, respectively. Spleen weights of the HFCD+D1.5 and HFCD+D7.5 groups were 0.15 ± 0.04 g and 0.10 ± 0.01 g, respectively. There were no significant differences between the groups after 13 weeks of their respective diets.
Fig. 1
Effects of PEITC and DIM supplementation on spleen weight and images of representative spleen tissues from mice fed a high fat/cholesterol diet. (A) Summary of spleen weight in each group. (B) Representative macroscopic images of dissected spleens from each group. There was no significant difference in spleen weights. Data presented as means ± SD (n = 10); different letters indicate significant differences (P < 0.05), as determined by applying Duncan's multiple range test. CON: AIN-93G diet, HFCD: 60% calories from fat+1% cholesterol, HFCD+P30: HFCD+30 mg/kg/day of PEITC, HFCD+P75: HFCD+75 mg/kg/day of PEITC, HFCD+D1.5: HFCD+1.5 mg/kg/day of DIM, HFCD+D7.5: HFCD+7.5 mg/kg/day of DIM.
PEITC, phenethyl isothiocyanate; HFCD, high fat/cholesterol diet; DIM, 3,3′-diindolylmethane.
Suppression of IL-6 secretion in serum by PEITC and DIM supplementation in HFCD-fed mice
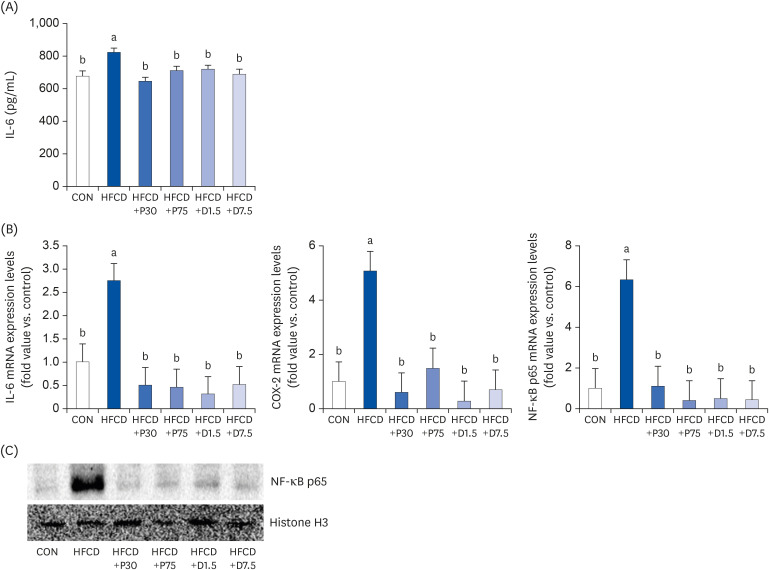
We measured serum IL-6 levels to identify HFCD-induced inflammation by using a commercial mouse IL-6 ELISA Kit. As shown in Fig. 2A, IL-6 secretion increased in the HFCD group compared to that in the CON group. However, the increase in IL-6 levels was suppressed in all of the HFCD+P and HFCD+D groups. These results indicate that the HFCD induced the secretion of pro-inflammatory cytokines in the serum. In addition, the HFCD-induced inflammation was modulated by administration of PEITC and DIM supplements, at least partially through a decreased secretion of IL-6.
Fig. 2
PEITC and DIM supplements suppress IL-6 secretion, IL-6 mRNA, COX-2 mRNA, and NF-κB p65 mRNA expressions and NF-κB p65 gene expression in serum and spleen of high fat/cholesterol diet-fed mice. (A) Pro-inflammatory IL-6 secretion was determined by enzyme-linked immunosorbent assay. Total RNA was subjected to qPCR analysis, as described in the materials and methods section. (B) Results of qPCR analysis of IL-6, COX-2, and NF-κB p65 gene expression in high fat/cholesterol diet-fed mice. The transcript mRNA levels of IL-6, COX-2, and NF-κB p65 were normalized to that of β-actin. (C) Lysates were prepared and underwent western blotting with NF-κB p65 antibody, as described in the materials and methods section. Increased NF-κB p65 expression was confirmed via western blot analysis. Values were analyzed using the 2-∆∆CT method. Significance was determined by comparison with β-actin normalized 2-∆∆CT values. Data presented as means ± SD (n = 3), and different letters indicate significant differences (P < 0.05, a > b), as determined by applying Duncan's multiple range test. CON: AIN-93G diet, HFCD: 60% calories from fat + 1% cholesterol, HFCD+P30: HFCD+30 mg/kg/day of PEITC, HFCD+P75: HFCD+75 mg/kg/day of PEITC, HFCD+D1.5: HFCD+1.5 mg/kg/day of DIM, HFCD+D7.5: HFCD+7.5 mg/kg/day of DIM.
IL-6, interleukin 6; COX-2, cyclooxygenase 2; PEITC, phenethyl isothiocyanate; DIM, 3,3′-diindolylmethane; HFCD, high fat/cholesterol diet; NF-κB, nuclear factor kappa B; qPCR, quantitative polymerase chain reaction.
Inhibition of inflammatory protein expressions in spleen by PEITC and DIM supplementation in HFCD-fed mice
We hypothesized that PEITC and DIM supplements could inhibit obesity-induced splenic inflammation in HFCD-fed mice. The qPCR results revealed that the transcription levels of COX-2, IL-6, and NF-κB p65 mRNAs were significantly higher in the HFCD group than in the CON group (Fig. 2B). Contrastingly, the transcription of IL-6, COX-2, and NF-κB p65 mRNAs significantly decreased in each of the PEITC and DIM supplement groups from that in the HFCD group (P < 0.05). Additionally, we examined the expression of NF-κB p65 using western blotting (Fig. 2C). The expression of NF-κB p65 was higher in the HFCD group than in the CON group. However, NF-κB p65 expression decreased in the HFCD+P75 and HFCD+D7.5 groups from that in the HFCD group. The inflammation inhibitory effects of PEITC and DIM were partly due to the suppression of NF-κB p65 expression in the spleen. Therefore, our results demonstrated that PEITC and DIM have an inhibitory effect on inflammation by diminishing the expressions of IL-6, COX-2, and NF-κB p65.
Inhibition of TLR2/4 and MyD88 mRNA expression levels in spleen by PEITC and DIM supplementation in HFCD-fed mice
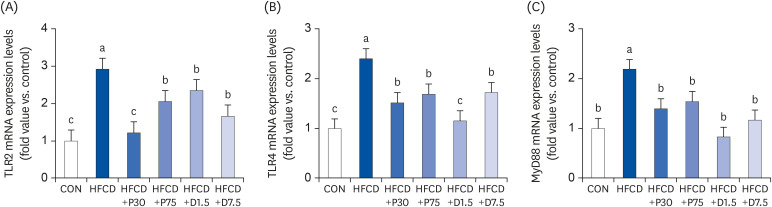
We also investigated the involvement of the TLR2/4-MyD88 pathway in the regulation of splenic inflammation by PEITC and DIM. TLR signals are negatively regulated to prevent or terminate excessive immune responses [7]. We measured the mRNA levels of TLR2, TLR4, and MyD88 in mouse spleen tissue of PEITC- and DIM-supplemented HFCD-fed mice (Fig. 3). The qPCR results revealed that TLR2, TLR4, and MyD88 mRNA expression levels were significantly higher in the HFCD group than in the CON group (P < 0.05). However, TLR2, TLR4, and MyD88 mRNA levels were lower in all HFCD+P and HFCD+D groups than that in the HFCD group (P < 0.05). These data suggest that PEITC and DIM supplementation might modulate inflammation triggered by immune processes.
Fig. 3
PEITC and DIM suppress (A) TLR2, (B) TLR4, and (C) MyD88 mRNA expressions in the spleen of high fat/cholesterol diet-fed mice. Results of qPCR analysis of TLR2, TLR4, and MyD88 gene expression in high fat/cholesterol diet-fed mice. Total RNA was subjected to qPCR analysis, as described in the materials and methods section. The transcript mRNA levels of TLR2, TLR4, and MyD88 were normalized to that of β-actin. Values were analyzed using the 2-∆∆CT method. Significance was determined by comparison with β-actin normalized 2-∆∆CT values. Data presented as means ± SD (n = 3), and different letters indicate significant differences (P < 0.05, a > b > c), as determined by applying Duncan's multiple range test. CON: AIN-93G diet, HFCD: 60% calories from fat+1% cholesterol, HFCD+P30: HFCD+30 mg/kg/day of PEITC, HFCD+P75: HFCD+75 mg/kg/day of PEITC, HFCD+D1.5: HFCD+1.5 mg/kg/day of DIM, HFCD+D7.5: HFCD+7.5 mg/kg/day of DIM.
TLR, Toll-like receptor; HFCD, high fat/cholesterol diet; MyD88, myeloid differentiation primary response 88; DIM, 3,3′-diindolylmethane; PEITC, phenethyl isothiocyanate; qPCR, quantitative polymerase chain reaction.
DISCUSSION
Increasing attention is being paid to the spleen due to its key roles in regulating the immune response [20] and dampening the inflammatory response [21]. Moreover, obesity is increasing rapidly worldwide, and diseases related to obesity are under focus [21]. It is widely reported that obesity can produce inflammatory conditions that are directly related to the onset of many diseases [22]. In addition, previous studies have indicated that obesity impairs immune function [23]. Although many studies have reported the physiological effects of PEITC and DIM, their effects on obesity-related inflammation of the spleen and the mechanisms underlying those effects have not been described.
The aim of this study was to examine the effects and functional mechanisms of PEITC and DIM in obesity-related splenic inflammation. A HFCD diet can induce the development of metabolic syndrome, which includes obesity, dyslipidemia, and a pro-inflammatory state [2425]. As mentioned, we previously observed that a HFCD increases body weight, adipose tissue weight, and liver tissue inflammation in mice over a 13-week study period [19].
Gotoh et al. [26] demonstrated that diet-induced obesity impaired the spleen's ability to synthesize pro-inflammatory cytokines in mice. Released from the spleen, these cytokines flow directly into the liver through the spleen and portal vein. As a result, the natural killer cytotoxicity level in the liver increases [26]. Despite the important functions of the spleen, the relationship between a HFCD and splenic inflammation risk factors in mice was unclear.
Thus, we examined spleen weight, immune processes, and pro-inflammatory markers in HFCD-fed mice. As shown in Fig. 1, we measured the spleen weight to observe the effect of obesity on the spleen. According to a study by Altunkaynak et al. [27], splenomegaly was observed in female Sprague Dawley rats fed a high-fat diet (HFD) for 3 months. Most studies have shown splenomegaly to be a result of HFD, but a few studies have shown the opposite. Trufakin et al. [28] reported that spleen weight decreases in HFD-fed Wistar rats. Such contradictory results can occur in different animal models. Interestingly, in this study, there was no difference in spleen weights among the various groups.
Kuo et al. [29] reported that curcumin caused a drastic reduction in the secretion of pro-inflammatory cytokines, including tumor necrosis factor-α, MCP-1, and IL-6 in the serum of obese mice. Consistent with that report, our data also observed that serum IL-6, which was increased in obese mice, was significantly reduced by PEITC and DIM supplementation.
Herein, we measured the effects of PEITC and DIM on splenic inflammation in mice fed a HFCD. We observed that PEITC and DIM significantly inhibited the expression levels of IL-6, COX-2, and NF-κB p65 in the spleen (Fig. 2). NF-κB is responsible for producing cytokines, chemokines, and growth factors, thereby regulating the expression of genes involved in immune and inflammatory responses [30]. Our results suggest that PEITC and DIM supplementation inhibits the inflammatory molecules COX-2, IL-6, and NF-κB p65 in splenic inflammation induced by a HFCD. Furthermore, TLR activates a pathway that leads to the activation of NF-κB p65 signaling [7].
However, there have been no reports on the links between the functions of PEITC and DIM and the activation of NF-κB p65 related to splenic inflammation in mice with HFCD-induced obesity. In our study, PEITC and DIM supplementation decreased TLR2/4, MyD88, and NF-κB p65 expression levels. We believe that these results indicate that PEITC and DIM supplementation can inhibit NF-κB p65 signaling, which is linked to TLR signaling via MyD88.
In this study, we conclude that PEITC and DIM diet supplements can alleviate inflammation of the spleen via suppression of TLR2/4 and MyD88 in obese mice. However, the exact mechanisms by which PEITC and DIM regulate these processes, especially in the spleen, need further investigation.
 XML Download
XML Download