

 PDF
PDF Citation
Citation Print
Print


INTRODUCTION
Oxysterols are oxygenated derivatives of cholesterol. As compared to cholesterol, they contain an additional hydroxy, epoxide or ketone group in the sterol nucleus, and/or a hydroxyl group in the side chain [1]. 27-Hydroxycholesterol (27OHChol) is a side-chain oxysterol oxygenated at the 27th carbon atom of cholesterol. This oxysterol is produced via oxidation by sterol 27-hydroxylase (CYP27A1), and metabolized via 7α-hydroxylation for bile acid synthesis in the liver [1,2]. 27OHChol can also be formed in peripheral tissues either via enzymatic or non-enzymatic mechanisms [2,3]. The production of 27OHChol is enhanced in atherosclerotic lesions of the artery, and increasing amounts detected in lesions are associated with increasing macrophages [4,5]. The biological function of 27OHChol in peripheral tissues differs from that in the liver. 27OHChol enhances inflammation and triggers an immune response by activating monocytes/macrophages. Following activation, they express chemokines, including chemokine (C-C motif) ligand (CCL) 2, CCL3 and chemokine (C-X-C motif) ligand (CXCL) 8, and pattern recognition receptors that respond to the bacterial pathogen-associated molecular patterns [6-8]. 27OHChol also induces molecules involved in T cell activation on the surface of monocytic cells [9]. Vascular smooth muscle cells and T cells, however, do not express such molecules in response to 27OHChol [10,11]. These findings indicate that macrophages are the major cell type whose gene expression is influenced by 27OHChol.
27OHChol is a functional liver X receptors (LXR) agonist [12]. LXRα (NR1H3) and LXRβ (NR1H2) are two isoforms of LXRs expressed with overlapping, but a distinctive pattern. LXRα is dominant in the liver and expressed primarily in the intestine, adipose tissue, and macrophages, whereas LXRβ is widely expressed [13-15]. Since LXRs are members of the nuclear hormone receptor superfamily of ligand-activated transcription factors, they exert their biological effects by controlling the expression of target genes [15]. Activation of LXRs with 27OHChol increases the transcription of LXR-responsive genes involved in cholesterol efflux, such as ABCA1 and ABCG1 in macrophages [12,14]. These findings indicate that 27OHChol induces gene expression via LXRs. However, it is unknown whether the LXR agonistic activity of 27OHChol is involved in the induction of genes other than for cholesterol metabolism.
This study was therefore undertaken to determine the effects of GSK 2033, a potent cell-active LXR antagonist, on the transcription of 27OHChol-induced genes, including inflammatory and cell surface molecules as well as LXR target genes. Our results demonstrate that GSK 2033 differentially regulates the 27OHChol-induced genes in macrophages.
METHODS
Cells and reagents
The human THP-1 monocyte/macrophage cell line (ATCC, #TIB-202) was purchased from the American Type Culture Collection (ATCC, Manassas, VA, USA). THP-1 cells were cultured in RPMI 1640 medium supplemented with 10% fetal bovine serum (FBS) at 37°C in a humidified atmosphere of 5% CO2. Penicillin (50 U/ml) and streptomycin (50 μg/ml) were added to prevent bacterial contamination. 27OHChol and antibodies against LXRα/β, CD14, CD80, and CD86 were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA). GSK 2033 and TO 901317 were purchased from Sigma-Aldrich (St. Louis, MO, USA). Anti-ABCA1 antibody was purchased from Invitrogen (Eugene, OR, USA). Anti-CD163 antibody conjugated with FITC and anti-CD206 antibody conjugated with PE were purchased from BioLegend (San Diego, CA, USA).
Reverse transcription-quantitative real-time polymerase chain reaction (RT-qPCR) analysis
Total RNA isolated were reverse-transcribed for 1 h at 42°C with 100 U Moloney murine leukemia virus reverse transcriptase in a 10 μl reaction volume, containing 50 mM Tris-HCl (pH 8.3 at 25°C), 55 mM KCl, 3 mM MgCl2, 10 mM DTT, 1 μg oligo (dT) 15 primers, 0.125 mM each dNTP, and 40 U RNase inhibitor. Subsequent qPCR was performed in triplicate using a LightCycler 96 Real-Time PCR System (Roche, Mannheim, Germany), as previously described [8]. Each 20 μl reaction mixture consisted of 10 μl SYBR Green Master Mix, 2 μl forward and reverse primers (10 pM each) of the gene to be quantified, and cDNA template. The thermal cycling conditions consisted of 95°C for 10 min, followed by 45 cycles of 95°C for 10 sec, 50°C for 10 sec, and 72°C for 10 sec. The relative expression of each gene was calculated as the ratio to the GAPDH gene using the LightCycler 96 software (Version 1.1.0.1320; Roche). The primers used were as follows in Table 1.
Western blot analysis
Cell lysates were separated by 10% SDS-PAGE, and resolved proteins were transferred to nitrocellulose membranes. After blocking for 1 h in 1% skim milk in TBS (pH 7.4) containing 0.05% Tween-20, membranes were incubated with antibodies against LXRα/β, ABCA1 or β-actin at 4°C overnight. Membranes were washed three times with 0.05% Tween 20/TBS for 10 min each and incubated for 1 h with HRP conjugated secondary Abs (1:5,000 dilution) at room temperature. After washing with 0.05% Tween 20/TBS, membranes were exposed to chemiluminescent detection reagents (Pierce ECL Western Blotting Substrate; Thermo Scientific, Rockford, IL, USA). Chemiluminescence images were captured by using an Amersham Imager 680 (GE Healthcare Life Scicences, Pittsburgh, PA, USA).
Flow cytometric analysis
THP-1 cells were harvested by centrifugation at 200 × g for 5 min at room temperature and incubated for 2 h with antibodies against CD14, CD80 or CD86 (1:100 dilution) in the FACS buffer containing 2 mM EDTA and 0.2% BSA in PBS. After washing with cold-PBS, cells were incubated for 1 h with Alexa Fluor 488-conjugated secondary antibodies (1:200 dilution) at 4°C. Or the harvested cells were incubated with the fluorescent dye-conjugated antibodies (1:100 dilution) against CD163 or CD206 for 1 h at 4°C. Cells were washed with cold-PBS, resuspended in 1% paraformaldehyde, and analyzed by flow cytometry.
ELISA
The amounts of CCL2, CCL3, CCL4, CXCL8, and TNF-α secreted in culture media were quantified using commercially available ELISA kits (R&D Systems, Minneapolis, MN, USA), following the manufacturer's instructions.
Statistical analysis
Statistical analysis was performed via one-way analysis of variance, followed by Dunnett's multiple comparison test, using PRISM (version 5.0) (GraphPad Software Inc., San Diego, CA, USA). A p-value less than 0.05 (p < 0.05) is considered to indicate a statistically significant difference.
RESULTS
GSK 2033 inhibits the transcription of LXR target genes
The human LXRα and ABCA1/ABCG1 are targets for regulation by LXR, and ligands for the receptor induce their expression in macrophages [12]. We investigated whether 27OHChol affects expression of the genes along with TO 901317, a synthetic LXR ligand. We observed levels of LXRα transcripts were elevated to 2.25- and 3.49-fold following stimulation with 27OHChol and TO 901317, respectively (Fig. 1A). Assessing the transcript levels of ABCA1 revealed increases up to 1.31- and 8.04-fold by stimulation with 27OHChol and TO 901317, respectively (Fig. 1B). ABCG1 transcripts increased 4.33- and 28.5-fold after stimulation with 27OHChol and TO 901317, respectively (Supplementary Fig. 1). Investigating further whether expressions of LXRα and ABCA1/ABCG1 genes were influenced by GSK 2033, a potent cell-active LXR antagonist [16], revealed that TO 901317- or 27OHChol-induced gene transcription of LXRα and ABCA1/ABCG1 was blocked in the presence of GSK 2033. In line with the results of real-time PCR, 27OHChol and TO 901317 elevated protein levels of LXRα or ABCA1, which was completely inhibited by GSK 2033 (Fig. 1C). These results indicate that although less effective than the synthetic LXR agonist, 27OHChol induces expression of the LXR target genes, and the induction is impaired by GSK 2033.
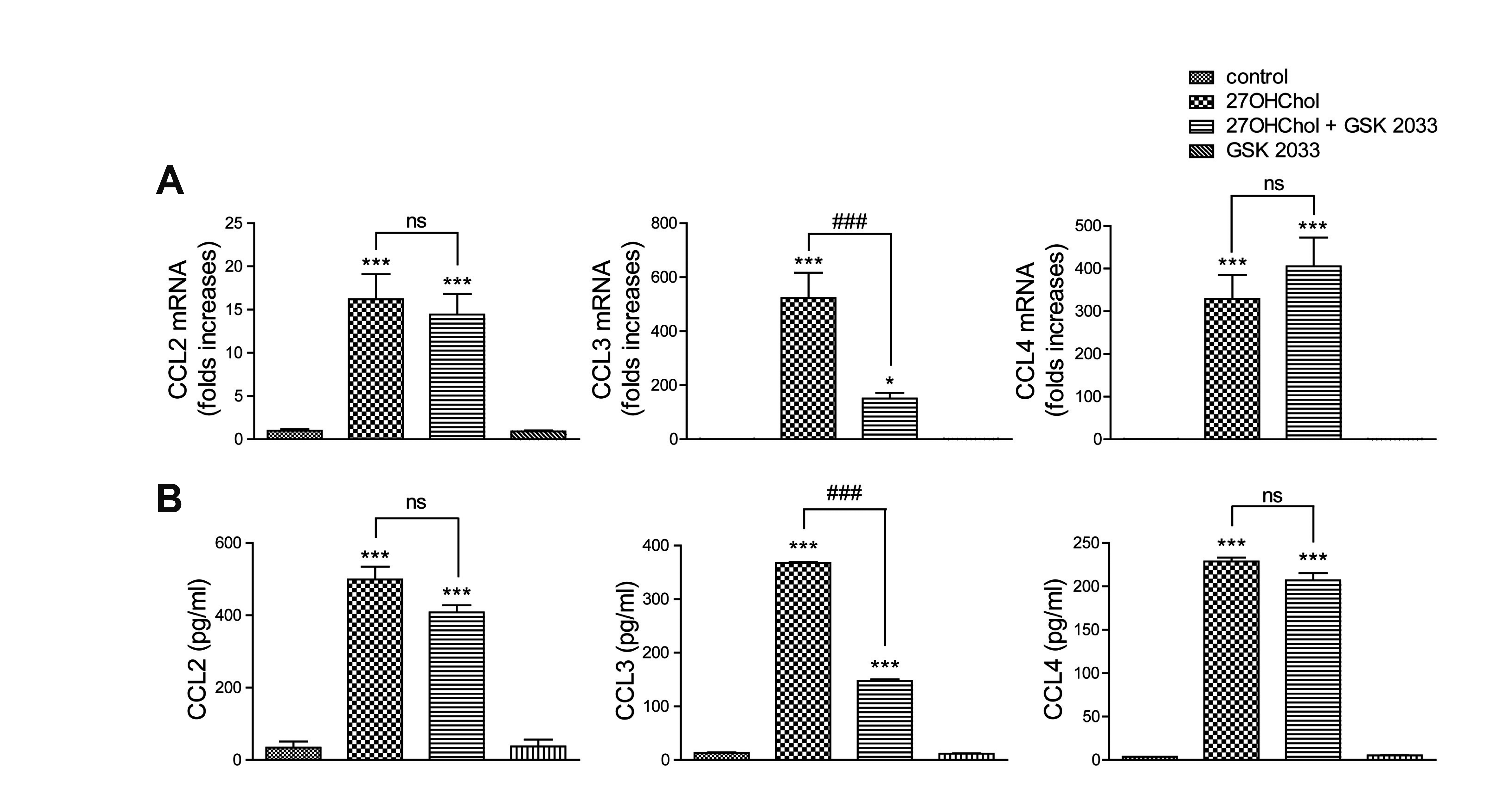
GSK 2033 suppresses 27OHChol-induced CCL3 gene
27OHChol induces expression of C-C chemokines, including CCL2, CCL3, and CCL4 [11]. We undertook to determine effects of GSK 2033 on the transcription of chemokines to understand the function of LXR inhibition in the induction of C-C chemokines. 27OHChol exposure elevated the levels of CCL2, CCL3, and CCL4 gene transcripts and their gene products (Fig. 2A and B). Although GSK 2033 did not significantly reduce expression of the CCL2 and CCL4 genes, the levels of CCL3 transcripts and protein were profoundly decreased in the presence of GSK 2033. The antagonist alone did not alter the transcript levels and secretion of the C-C chemokines. These results suggest that of the three C-C chemokines, GSK 2033 suppresses expression of the CCL3 gene alone.
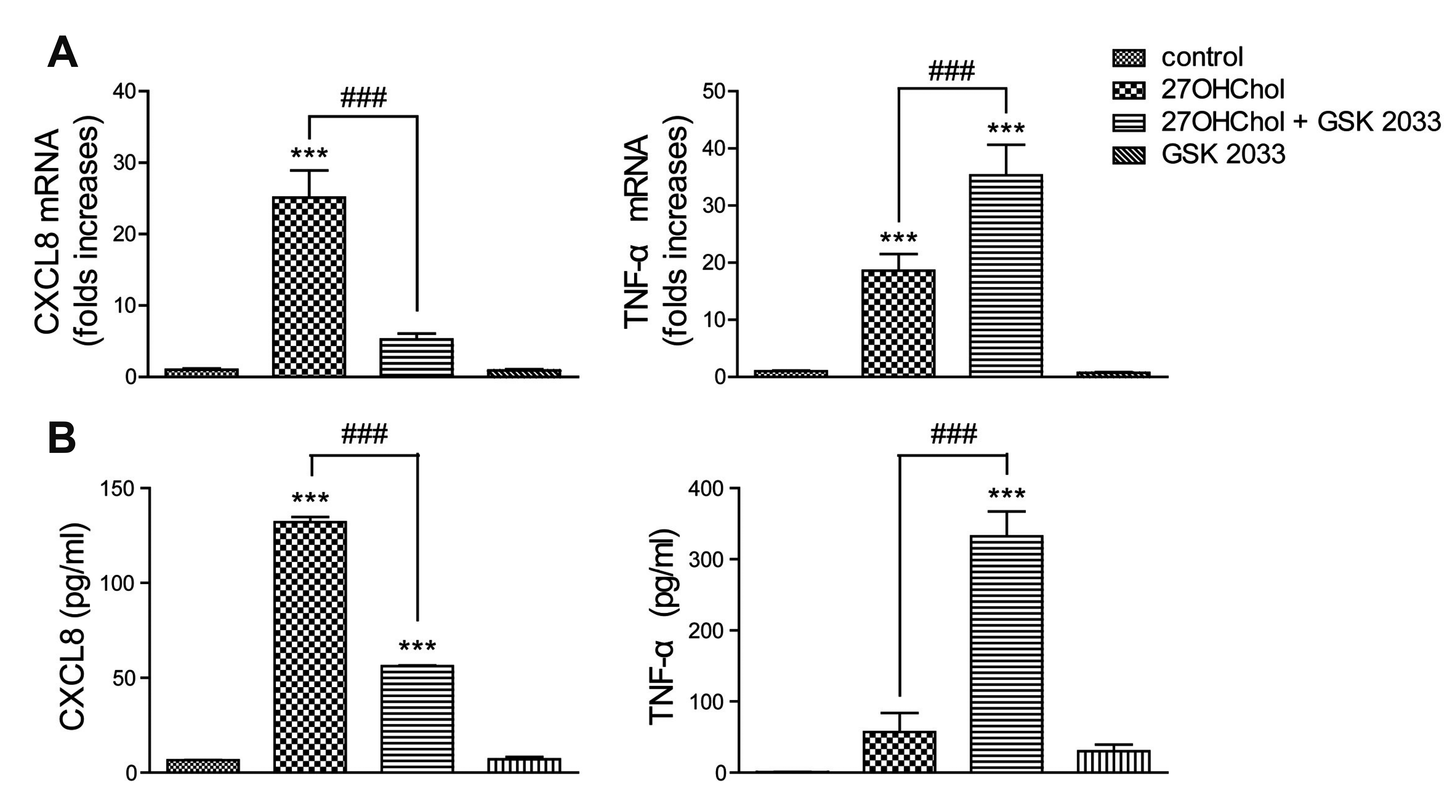
GSK 2033 displays opposite effects on 27OHChol-induced CXCL8 and TNF-α
27OHChol as well as LXR agonists induces the CXCL8 and TNF-α genes which are involved in acute-phase inflammatory responses [8,17,18]. We investigated the roles of LXR agonistic activity of 27OHChol in the expression of CXCL8 and TNF-α. Treatment with GSK 2033 resulted in suppressing the transcription of CXCL8 gene, whereas the transcription of TNF-α further enhanced in the presence of GSK 2033 (Fig. 3A). We examined effects of GSK 2033 on CXCL8 and TNF-α secretion. GSK 2033 impaired CXCL8 production, but promoted secretion of TNF-α (Fig. 3B). These results suggest that LXR-agonistic activity of 27OHChol is associated with a repressive effect on TNF-α expression.
GSK 2033 downregulates the transcription of M1/M2 surface markers
27OHChol affect levels of both M1 and M2 markers of macrophages [8]. We used GSK 2033 to estimate involvement of LXR agonistic activity in the induction of M1/M2 markers. 27OHChol increased levels of transcripts and surface expression of M2 markers of CD163 and CD206. Exposure to GSK 2033 resulted in suppressing the transcription of CD163 and CD206 genes and downregulating their surface levels (Fig. 4A and B). We also investigated effects of GSK 2033 on M1 markers of CD80 and CD86, along with CD14. The transcription of three genes elevated by 27OHChol was suppressed in the presence of GSK 2033. The LXR antagonist, however, did not downregulate surface expression of CD14, CD80, and CD86 (Fig. 5A and B). These results suggest that LXR activation is required for 27OHChol to induce transcription of the M1/M2 markers and that they are differentially regulated at protein level after LXR inhibition.
DISCUSSION
The LXRs are members of the nuclear receptor superfamily that modulate metabolism, development, proliferation and inflammation through positive and negative regulation of gene expression [13]. LXRs have crucial functions in the regulation of immune responses. Activation of LXR with agonists downregulates the expression of inflammatory genes through a process known as transrepression, and several studies have demonstrated the anti-inflammatory activities of synthetic LXR agonists using different mouse models of inflammatory diseases [19-23]. Endogenous oxysterols, like 27OHChol, are known to activate LXRs [12]. It is evident that this oxysterol is a pro-inflammatory molecule activating macrophages. Activated cells increase cytokine production and upregulate cell surface molecules involved in the immune response [7,9,10]. The current study investigated whether LXR-agonistic activity of 27OHChol affects the cytokine production and upregulates surface molecules after exposure to cell-active GSK 2033 [16], and thereby explored the biological functions of LXRs in macrophage activation.
27OHChol induces expression of diverse genes involved in lipid metabolism, inflammation, and cell differentiation [9,10,12]. Consistent with previous reports [12,14], we found that 27OHChol induces the LXR target genes of LXRα and ABCA1/ABCG1 via LXR-dependent mechanisms. Besides, we have demonstrated that the LXR agonistic activity is involved in expression of CCL3, CXCL8, CD14, CD80, CD86, CD163 and CD206, since their gene transcription is inhibited by GSK 2033. The significantly reduced expression of pattern recognition receptors, such as CD14 and CD206, following LXR inhibition, agrees with previous study that has demonstrated that LXR activation impacts the functioning of immune cells to establish an immune response against pathogens [24-26]. The downregulation of CD80 and CD86 is in line with a previous study by Torocsik et al. that reported increased expression of these molecules in monocyte-derived dendritic cells after administration of LXR agonists [27]. Taken together, our results suggest that the LXR-agonistic activity of 27OHChol is involved in the complex effects of this oxysterol on immune cells.
27OHChol elevates transcripts of the LXRα gene without a noticeable increase in its gene product. TO 901317, however, enhances its expression at both mRNA and protein levels, as shown in Fig. 1. The differences can be caused by the nature of LXR agonistic activity. 27OHChol is a natural LXR agonist, whereas TO 901317 is a synthetic one. TO 901317 displays a strong LXR agonistic activity and fully activates LXR signaling pathways [23]. In contrast, 27OHChol has a weak partial LXR agonistic activity [12]. Even though 27OHChol increases the level of LXRα transcripts, it seems that the activation of additional factors is required for translation of the mRNA into protein.
Cell surface molecules such as CD14, CD80, CD86, CD163, and CD206 are specific markers whose levels are enhanced by 27OHChol on monocytic cells [8]. Among them, CD163 and CD206 belong to M2 polarization markers, and CD80, and CD86 to M1 markers [28]. The results of Figs. 4B and 5B imply that expression of CD163 and CD206 protein is positively regulated by the LXR pathway while CD14, CD80, and CD86 are negatively regulated at protein level in the presence of 27OHChol. Collectively, these results suggest that LXR activity of 27OHChol is also involved in M2 polarization, which agrees with a previous study that reported that 27OHChol drives M2 polarization of human macrophages [29].
In summary, LXR inhibition with GSK 2033 leads to inhibited expression of CCL3 and CXCL8, but not of CCL2 and CCL4, enhanced TNF-α expression, and differential expression of M1/M2 markers on cell surface. We believe that the diverse effects of GSK 2033 are likely to be specific. GSK 2033 alone did not affect cell viability (Supplementary Fig. 2) and had no effect on the basal expression of the aforementioned genes. These results indicate that 27OHChol activates macrophages and affects immune cells via LXR-dependent and -independent mechanisms. Since 27OHChol induces inflammation and modifies immune responses by activating primarily monocytic cells, the molecular mechanisms underlying the cell type-specific effects of this oxysterol are still subject to further studies.
SUPPLEMENTARY MATERIALS
Supplementary data including two figures can be found with this article online at https://doi.org/10.4196/kjpp.2021.25.2.111.
 XML Download
XML Download