

 PDF
PDF Citation
Citation Print
Print


INTRODUCTION
Carbon monoxide (CO), a biological gas, is known to have highly toxic and detrimental effects on the heart [1,2]. CO exposure can induce arrhythmia [3,4] and myocardial cell death, leading to cardiac fibrosis [5]. However, CO is now established as an important, biologically active signaling molecule generated through the heme oxygenase (HO)-catalyzed degradation of heme [6,7]. Atrial and ventricular cardiomyocytes constitutively express HO-2, and various stress factors, including myocardial infarction [8], can increase the levels of inducible HO-1 [9]. Endogenously synthesized CO is being increasingly recognized as a potential therapeutic with important signaling functions in various diseases [10]. HO-derived CO protects the heart from transplant-associated ischemia-reperfusion injury [11]. The remarkable cardioprotective effects of HO-1 are best evidenced by its ability to regulate inflammatory processes, cellular signaling, and mitochondrial function, ultimately mitigating myocardial tissue injury and the progression of vascular proliferative disease [7].
Cardiac fibroblasts are the largest cell population in the permanent cellular constituents of the heart, which include cardiomyocytes, endothelial cells, and vascular smooth muscle cells [12]. Human cardiac fibroblasts (HCFs) have numerous functions, including the synthesis and deposition of extracellular matrix, and they play a relevant role in myocardial structuring and cell signaling in healthy and diseased myocardium [13]. HCFs have cell-cell communication with cardiomyocytes and other cells [14], and the cardiomyocyte–cardiac fibroblast interactions are important in normal heart function and in the development of diseases such as cardiac arrhythmia and fibrosis [15].
It has been reported that cardiac fibroblasts can interact electrically with cardiomyocytes through gap junctions [16] and direct electrical coupling of these two types of cells has also been observed [16-18]. There is now increasing evidence that cardiac fibroblasts may play a direct role in modulating the electrophysiological substrate in healthy and diseased hearts [19]. In addition, cardiac injury results in significant electrophysiological changes that enhance fibroblast-myocyte interactions and could contribute to a greater incidence of arrhythmias observed in fibrotic hearts [20].
Although cardiac fibroblasts are non-excitable, they express multiple ion channels and the activity of ion channels in HCFs [21,22] contributes to the functional activities of heart cells through the transfer of electrical signals between these two cell types [23]. However, the distribution and properties of their ion channels are quite distinct from those of cardiomyocytes [24].
The large-conductance Ca2+-activated K+ (BK) channel is the main K+ channel in HCFs [22,25]. The BK channel contributes to the resting membrane potential of cardiac fibroblasts [26] and the electrical coupling of cardiomyocytes-fibroblasts [23]. BK channels are also mainly expressed in vascular smooth muscle cells [27] and in the inner mitochondrial membranes of the cardiomyocytes [28]. Activation of these channels in these locations results in cardioprotection against cardiac ischemia that induces arrhythmogenesis [29].
CO is rapidly emerging as an important cellular messenger, regulating a wide range of physiological processes. The investigation of ion channels as effectors of CO signaling is in its infancy, with regard to both the physiologic and the toxic activities of this gas. Various ion channels have recently been discovered to be effectors of CO signaling, and they play key roles in the mediation of beneficial effects of CO [30,31]. CO also modulates various ion channels via diverse signaling pathways [2,32,33].
Among them, CO activates BK channels in human endothelial cells directly as well as via a cGMP-dependent pathway [34] and in vascular smooth muscle cells directly but not mediated by a cGMP dependent pathway [35].
However, the effect of CO on the BK channel of HCFs and the underlying mechanism remains unclear. Therefore, we explored the effect of CO, using CORMs, on BK current through the channels and their intracellular signaling pathways.
METHODS
Cell culture and reagents
Adult human cardiac ventricular fibroblasts were obtained from the ScienCell Research Laboratory (Cat #6310; San Diego, CA, USA). The cells were cultured in Dulbecco’s modified Eagle’s medium (Welgene, Gyeongsan, Korea) with 10% fetal bovine serum (Welgene) and a penicillin-streptomycin solution (100×; Welgene) in an incubator with a humidified atmosphere of 5% CO2 and 95% air at 37°C. Experiments were performed with cells from passage 4–7 (passage is the number of times the cells are processed with trypsin and transferred to another flask).
CO was applied to cells using the commercially available CO-donors, carbon monoxide releasing molecules; CORM-2 (tricarbonyldichlororuthenium [II] dimer, [Ru(CO3)Cl2]2), CORM-3 (tricarbonylchloro‐glycinate‐ruthenium [II], [Ru(CO)3Cl‐glycinate]), paxilline (a BK channel blocker), and all other chemicals were purchased from Sigma-Aldrich (St. Louis, MO, USA).
Electrophysiological recordings
Membrane ionic currents were recorded using the whole-cell patch-clamp technique, as described previously, using the Axopatch 200B Patch Clamp Amplifier (Axon Instruments, Union City, CA, USA).
The recording patch pipettes were prepared from filament-containing borosilicate tubes (TW150F-4; World Precision Instruments, Sarasota, FL, USA) using a 2-stage microelectrode puller (PC-10; Narishige, Tokyo, Japan) and were fire-polished using a microforge (MF-830; Narishige).
The pipettes for whole-cell currents exhibited a resistance of 2–3 MΩ when filled with the internal pipette solution. The recorded membrane currents were filtered at 2 kHz and digitized at 10 kHz. pCLAMP 9.0 software (Axon Instruments) was used for data acquisition and analysis of the whole-cell currents. All electrophysiological experiments were performed at room temperature.
For BK current recording, the cells were perfused with Tyrode solution containing 142 mM NaCl, 5 mM KCl, 1 mM CaCl2, 1 mM MgCl2, 5 mM glucose, and 5 mM HEPES (pH-adjusted to 7.35 with NaOH). The pipette solution contained 145 mM KCl, 1.652 mM CaCl2 (pCa 6.0), 1.013 mM MgCl2, 10 mM HEPES, 2 mM EGTA, and 2 mM K-ATP (pH 7.3 with KOH). All chemicals were purchased from Sigma-Aldrich. To record only IBK in the cells, we added 4-aminopyridine (1 mM) into the bath solution to exclude the influence of delayed rectifier K+ channels, which are another source of the prominent K+ currents in HCFs.
Statistical analysis
The results are presented as means ± standard errors of the mean (SEM). Statistical analysis was performed using SPSS version 22.0 software (IBM Corp., Armonk, NY, USA). The paired Student’s t-test was used to evaluate differences between the means of the 2 groups, whereas one-way analysis of variance was used for multiple groups. The p-values < 0.05 were considered statistically significant.
RESULTS
Effects of CO on large-conductance Ca2+-activated K+ currents of HCFs
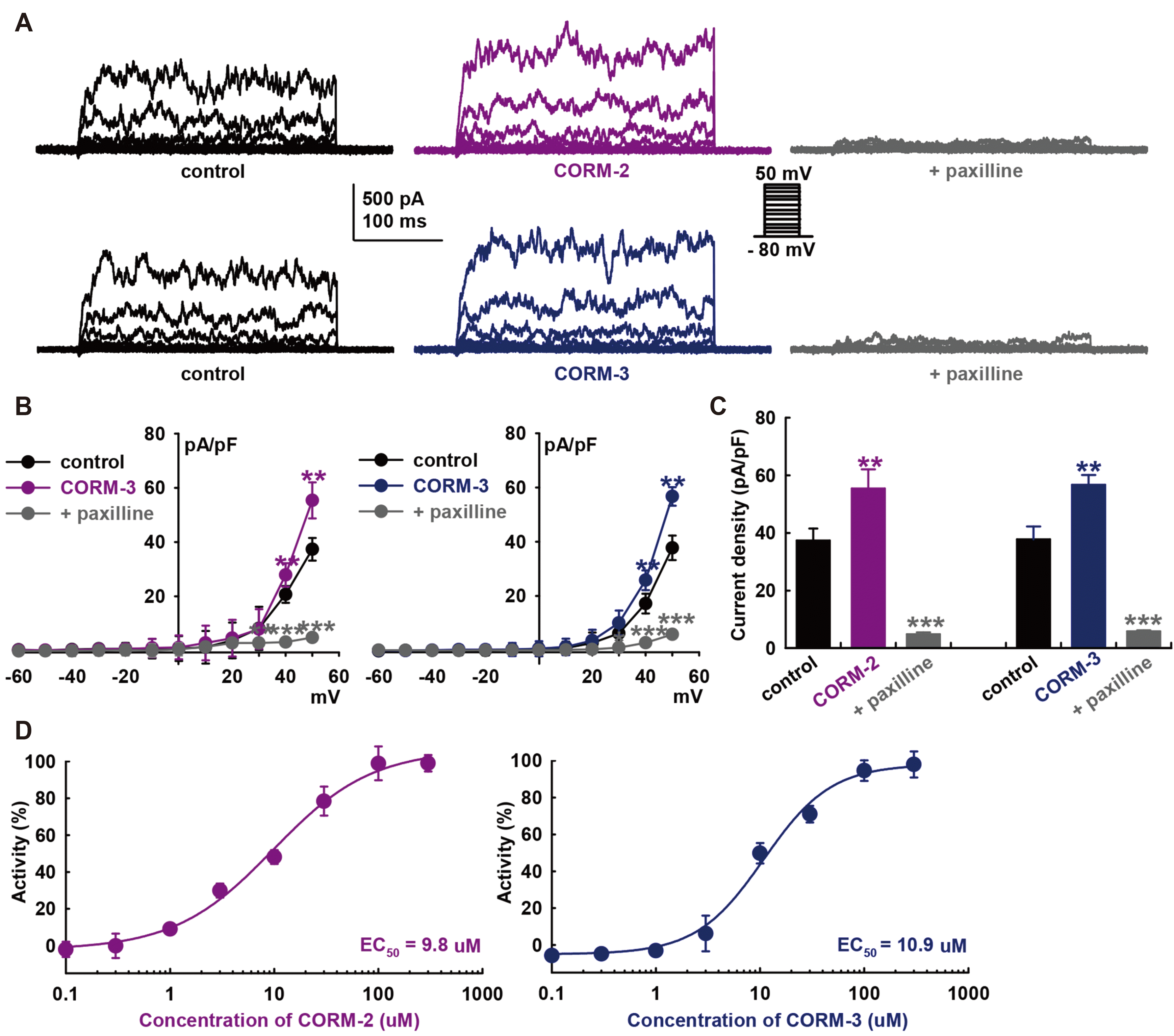
To determine the effect of CO on the BK channels in HCFs, we used whole-cell mode patch clamp recordings with a voltage protocol that consisted of depolarizing steps (from −80 mV to +50 mV) in 10-mV increments for 400-ms with a holding potential of −80 mV. The recorded macroscopic K+ currents of HCFs exhibit behaviors typical of BK currents (IBK): activated at 10 mV, increased in voltage-dependent manner, and strongly oscillated in response to strong depolarization, well maintained throughout the test pulse without marked inactivation during depolarizing voltage increments (Fig. 1A). The average cell capacitance was 23.44 ± 0.46 pF (n = 289). CO donors significantly increased the amplitude of the K+ currents (10 μM CORM-2, +48.2 ± 16.0% of control; CORM-3, +50.1 ± 7.6% of control, at +50 mV, n = 10, each, p < 0.01).
We then added 10 μM paxilline, a specific BK channel blocker, to confirm IBK and that the currents were blocked (CORM-2, –87.0 ± 1.6% of control; CORM-3, –84.4 ± 3.3 % of control, at +50 mV, n = 10, p < 0.001). In addition, their current–voltage (I–V) curves showed strong outward rectification, a characteristic of IBK (Fig. 1B). The bar graphs show the summary of the current density changes of these currents at +50 mV stimulation voltage (CORM-2, from 37.4 ± 4.2 to 55.4 ± 6.7 pA/pF; CORM-3, from 37.8 ± 4.5 to 56.7 ± 3.4 pA/pF; at +50 mV, n = 10, p < 0.01, Fig. 1C). Paxilline (10 μM) significantly inhibited CO-induced IBK activation (CORM-2, 4.9 ± 0.7; CORM-3, 5.9 ± 0.3 pA/pF, at +50 mV, n = 10, p < 0.001). Concentration–response curves of the CO donors showed steady-state currents normalized to the control data were fitted with the Hill equation (Fig. 1D), with the half maximal effective concentration (EC50) value of 9.8 μM for CORM-2 activation of IBK (EC50 of CORM-3; 10.9 μM).
Effects of NOS blockers on CO-induced IBK activation
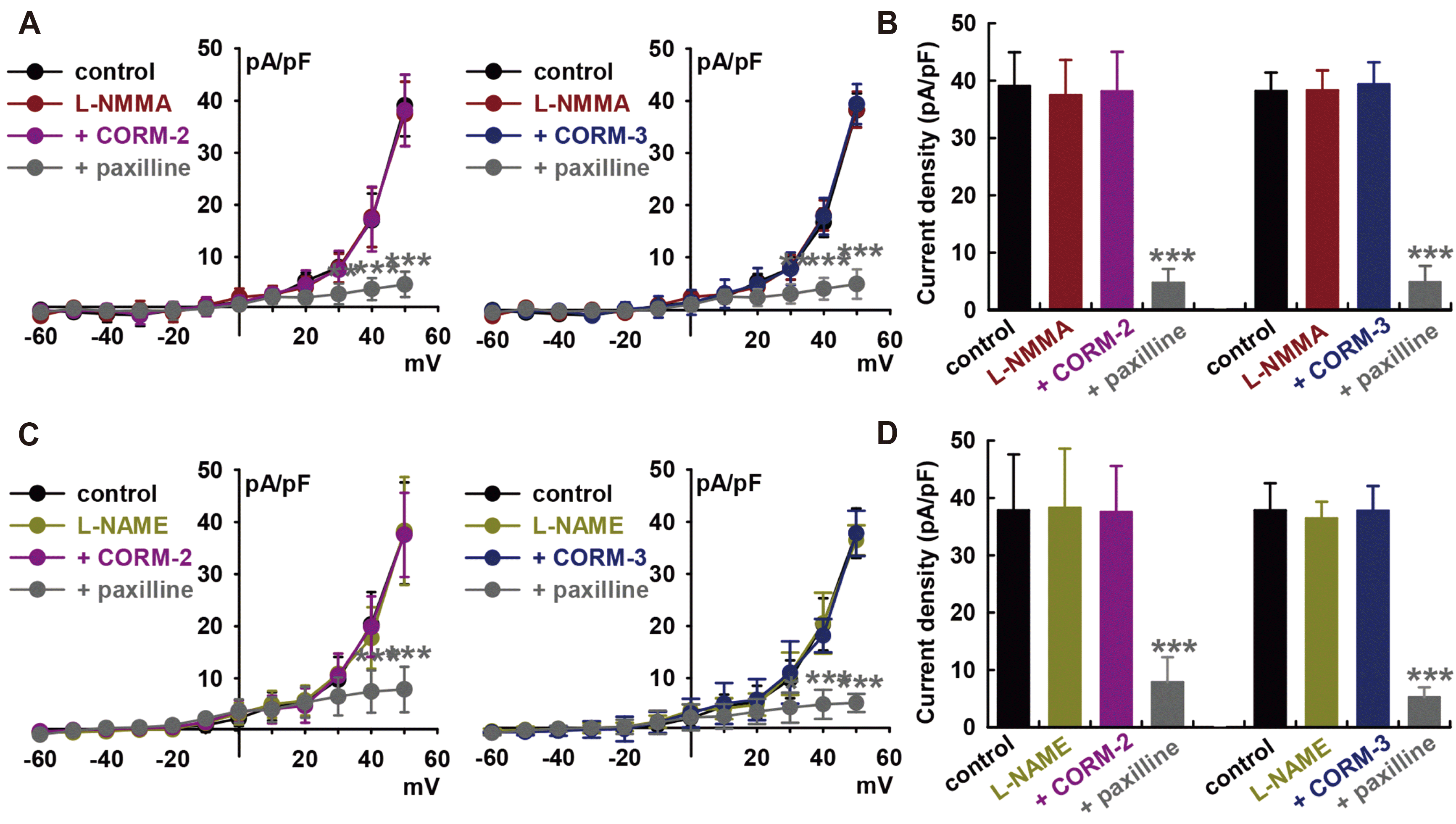
CO can regulate ion channels via the modulation of numerous signaling pathways. To investigate the signaling mechanism underlying the regulation of BK channels by CO, we first explored the involvement of nitric oxide (NO) because CO is known to activate nitric oxide synthase (NOS) and soluble guanylate cyclase (sGC) [36], and crosstalk between CO and NO have also been reported [37]. Fig. 2 indicates that the ability of CORM-2 (10 μM) to activate IBK was significantly suppressed by pre-treatment with an NOS inhibitor, L-NG-monomethyl arginine citrate (L-NMMA, 100 μM, –2.4 ± 11.7% of control, Fig. 2A and B). L-NMMA by itself did not affect the amplitude of IBK (–4.1 ± 10.4% of control, n = 10). Similarly, L-NMMA pre-treatment inhibited CORM-3-induced IBK activation (10 μM, +3.1 ± 11.9% of control, n = 10).
IBK was confirmed, which was significantly attenuated by 10 μM paxilline (CORM-2; –87.9 ± 4.1% of control, CORM-3; –87.3 ± 8.8% of control, n = 12, p < 0.001). Pre-treatment of cells with another NOS blocker, L-NG-nitroarginine methyl ester (L-NAME, 100 μM), also attenuated the CO-induced IBK activation. After 20 min of L-NAME pre-treatment, CORM-2 (–0.8 ± 8.3% of control, n = 12, Fig. 2C and D) or CORM-3 (–0.2 ± 9.1% of control, n = 12) could not increase IBK, and successive addition of paxilline after CO donors blocked the currents (CORM-2, –79.2 ± 4.5% of control; CORM-3, –86.1 ± 3.6% of control, p < 0.001). These data suggest that CO could activate IBK through NO formation via NOS.
Effect of cGMP signaling pathway on CO-induced IBK activation in HCFs
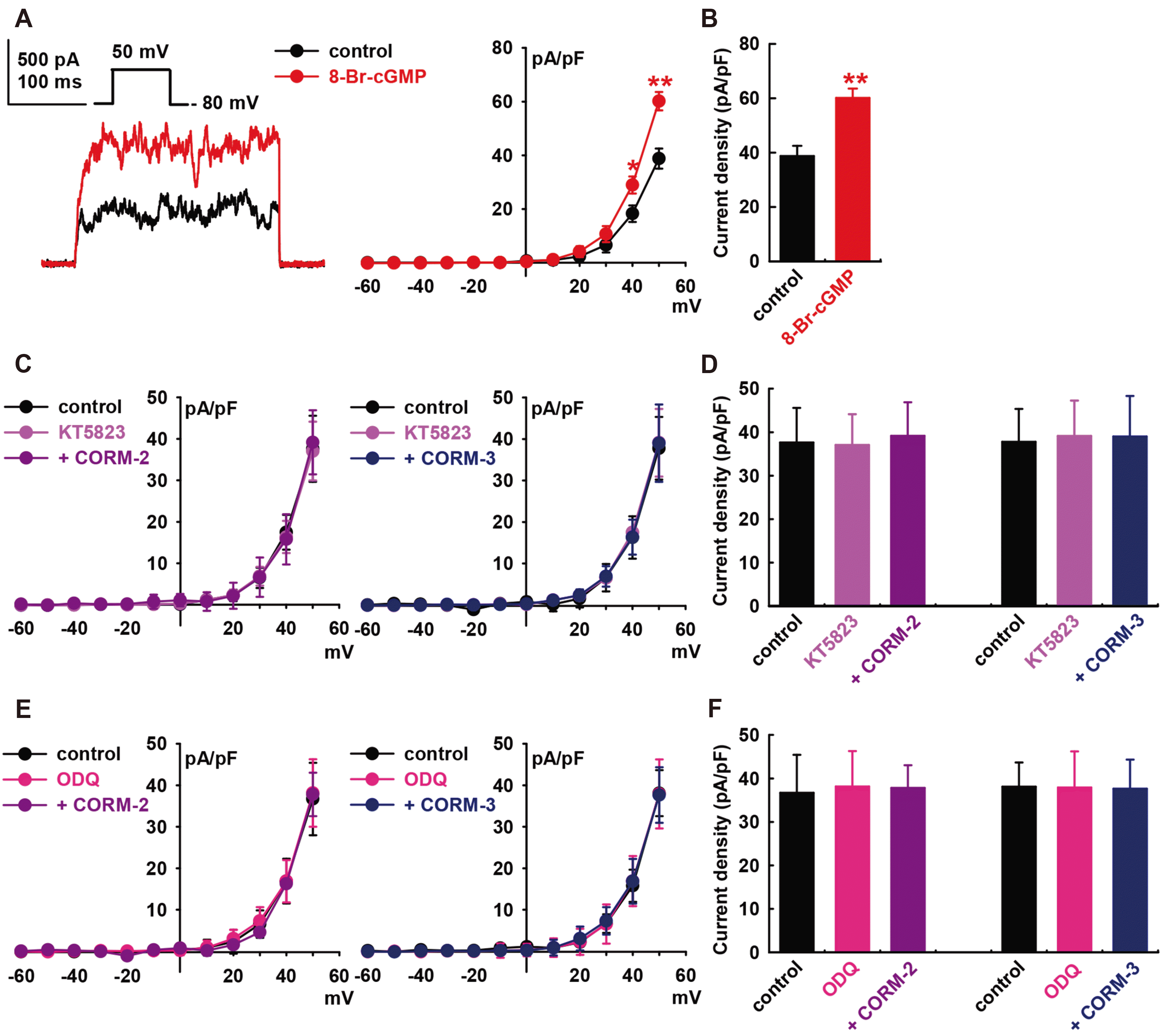
Binding of NO to the heme group of sGC leads to increased conversion of GTP to cGMP, which in turn activates PKG. The cGMP signaling pathway are the main mechanisms for mediating the effects of NO on IBK in HCFs [25]. Therefore, we assessed the effects of cGMP on IBK of HCFs. Addition of 8-bromo-cGMP (a membrane-permeable cGMP analogue, 300 μM) increased the IBK to +55.1 ± 9.2% of the control at +50 mV (p < 0.01, n = 8, Fig. 3A and B).
When we pre-treated the cells with a PKG inhibitor, KT5823 (1 μM), the CO donors failed to increase IBK (CORM-2, +4.1 ± 9.7% of the control at +50 mV, n = 8; CORM-3, +3.3 ± 12.3% of control, n = 8, Fig. 3C and D). Similarly, pre-treatment of the cells with 1H-[1,-2,-4] oxadiazolo-[4,-3-a] quinoxalin-1-one (ODQ; 10 μM, a membrane-permeable sGC inhibitor) inhibited the CO-induced activation of IBK (CORM-2, +3.1 ± 6.0% of the control at +50 mV, n = 8; CORM-3, –1.2 ± 12.1% of control, n = 8, Fig. 3E and F). KT5823 (1 μM) or ODQ (10 μM) alone did not increase the IBK of HCFs.
Effect of cAMP signaling pathways on CO-induced IBK activation of HCFs
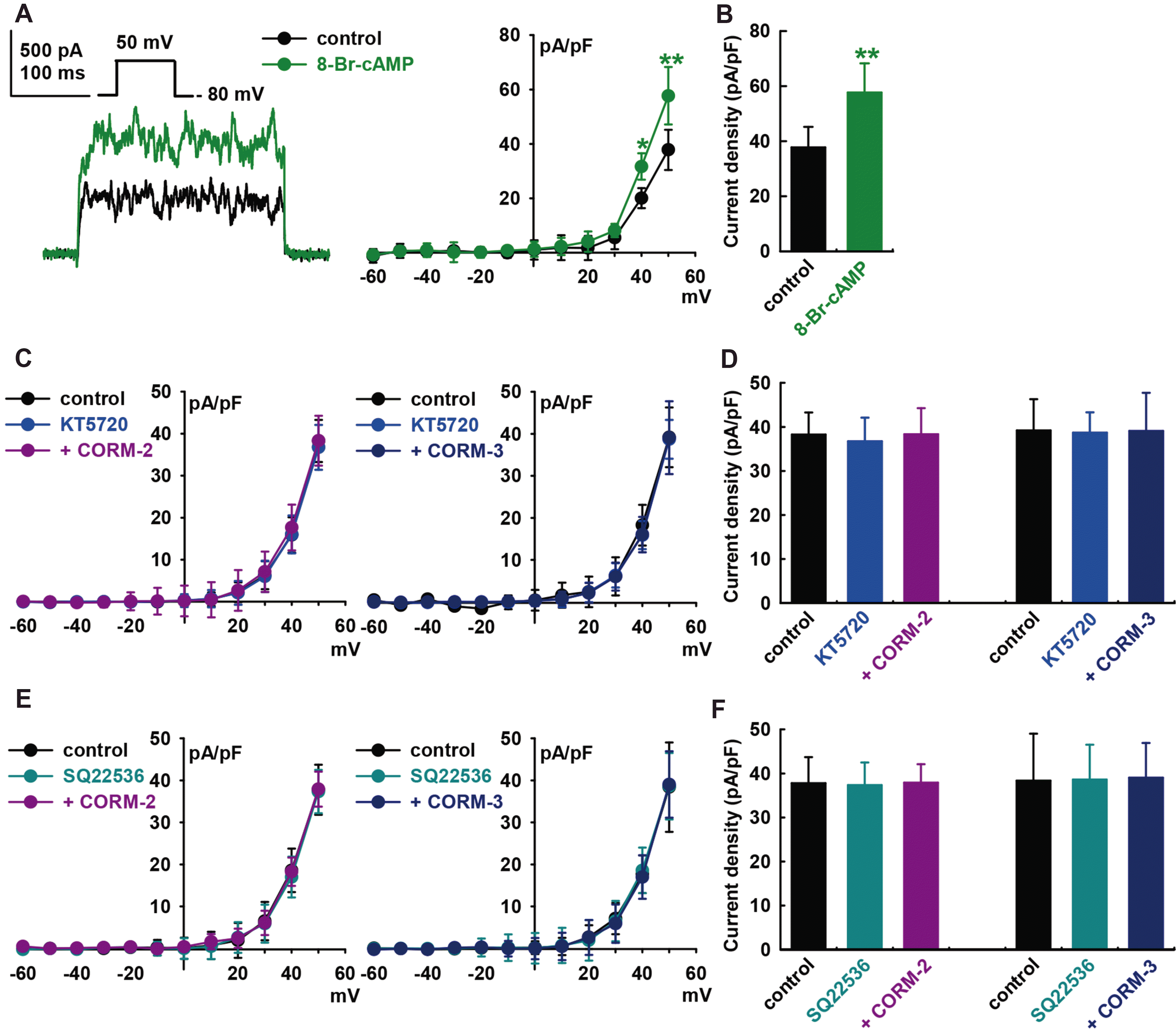
To determine whether the cAMP signaling pathways are also involved in CO-induced IBK activation, 8-bromo-cyclic AMP (8-Br-cAMP, a cyclic AMP analog, 300 μM) was added into the bath solution, which increased IBK to +52.7 ± 14.3% of the control at +50 mV (p < 0.01, n = 8, Fig. 4A and B). Pre-treatment of the cells with KT5720 (1 μM, a PKA blocker) for 20 min blocked CO-induced IBK activation (CORM-2, +0.2 ± 11.9% of the control, at +50 mV, n = 8; CORM-3, –0.2 ± 12.2% of the control, n = 8, Fig. 4C and D). Pre-treatment with SQ22536 (1 μM, an adenylate cyclase blocker) also inhibited CO-induced IBK activation (CORM-2, control, +0.4 ± 7.0% of the control, n = 8; CORM-3, +1.7 ± 7.4% of the control, n = 8, Fig. 4E and F). KT5720 (1 μM) or SQ22536 (1 μM) alone did not increase the IBK of HCFs.
Influence of S-nitrosylation on CO-induced IBK activation
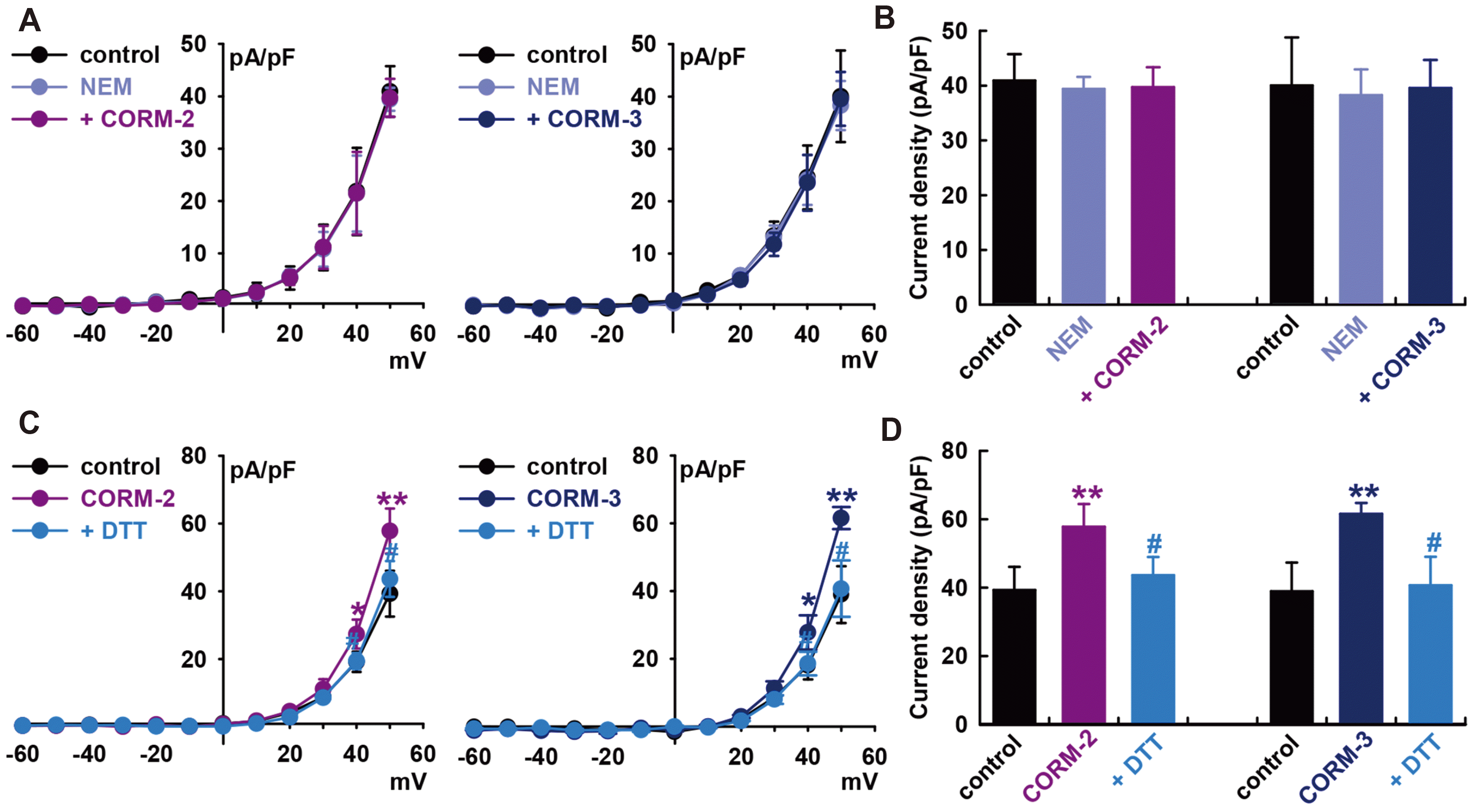
To establish S-nitrosylation as a mechanism for CO-mediated activation of IBK, we pre-treated cells with a thiol-alkylating reagent, N-ethylmaleimide (0.5 mM, NEM), for 20 min and then applied the CO donors. In the presence of NEM, CO donors could not increase IBK compared to the value in the presence of NEM alone (CORM-2, –3.0 ± 7.6% of the control, n = 6; CORM-3, –1.2 ± 5.9% of the control, n = 6, Fig. 5A and B).
When DL-dithiothreitol (DTT, a reducing agent; 5 mM) was applied after IBK had been increased by CORM-2, it reversed the effect of CORM-2 on IBK (CORM-2, +47.0 ± 9.8% of the control, n = 6, **p < 0.01 vs. control; DTT, +11.0 ± 7.8% of the control, n = 6, #p < 0.05 vs. CORM-2, Fig. 5C and D). In case of CORM-3, DTT also reversed the CORM-3-induced increase of IBK (CORM-3, +57.9 ± 3.9% of the control, n = 6, **p < 0.01 vs. control; DTT, +4.5 ± 9.9% of the control, n = 6, #p < 0.05 vs. CORM-3). These findings further suggest that S-nitrosylation is one of the mechanisms behind CO-induced IBK activation in HCFs, and a thiol residue could be the ultimate target of CO.
DISCUSSION
CO activation of the BK currents of HCF
In our results, CO produced a concentration-dependent activation of IBK (Fig. 1). This result is consistent with the results for BK channels in human endothelial cells [34], in vascular smooth muscle cells [35], and in the mitochondrial BK channels of cardiomyocytes [38].
Previously, we have shown that the presence of the BK channel in the plasma membrane of HCFs by RT-PCR and Western blotting [21] and here confirmed its presence functionally by applying an electrophysiological method with paxilline, a specific BK channel blocker that exclusively uses a closed-channel block mechanism [39]. BK channels of the plasma membrane share multiple biophysical similarities with the BK channels in the inner mitochondrial membrane [38]. The protective effects of mitochondrial BK channel activation against ischemia were demonstrated by using BK channel openers [40] and BK channel knockout mice [41]. Therefore, considering the electrical coupling of cardiomyocytes-fibroblasts [23], BK channels of the plasma membrane of HCF could also be a potential target for cardiovascular diseases [42], and CO as a BK channel activator could be employed as a cardioprotectant.
For studies on CO signaling, cells and channels have been exposed to CO by the application of CO-releasing molecules (CORMs) that are a group of compounds capable of carrying and liberating controlled quantities of CO into cellular systems [6]. CORMs are fully water soluble, allow for intravenous administration, and rapidly liberate CO and hence have been used as CO donors to overcome the limitations of using CO gas [43]. In addition, CORMs are valuable experimental tools and potential therapeutic agents [6]. They have the potential for vasodilatory, anti-ischemic, and anti-inflammatory effects [44,45] and they could protect adult cardiomyocytes against hypoxia-reoxygenation [46]. Therefore, the use of CORMs to investigate the signaling properties of CO has provided many new applications and treatments as pharmacologic approaches to cardiovascular diseases [6].
However, some of their actions can occur independently of CO release [47] or they show different activities. CO has positive inotropic activity in the perfused rat heart by CORM-3 but not by CORM-2 [48]. Judicial use of appropriate control compounds, as well as a comparison of their effects with those of CO diluted directly into a solution, should be performed wherever experimentally possible. When we tested two frequently used two types of CORMs of different structures to confirm the CO effect on BK channels, CORM-2 and CORM-3 showed similar activating effects on IBK of HCFs; the EC50 value was 9.8 μM for CORM-2 and 10.9 μM for CORM-3.
CO activation of BK currents of HCF through NO
To investigate the mechanism of the regulation of BK channels by CO, we first explored the involvement of NO because CO and NO are two endogenously produced gases that can act as second messenger molecules and it is becoming increasingly clear that these two gases do not always work independently, but rather can modulate each other's activity [37]. CO induces NO release [49] and NO increases the expression of HO-1 in endothelial cells [50] or vascular smooth muscle cells [51].
Our results also demonstrated that the activation of IBK by CO in HCFs was abolished by treatment with L-NMMA or L-NAME, NOS blockers (Fig. 2). These results are consistent with the finding that CO activates L-type calcium channels in HEK cells and in human intestinal smooth muscle cells [36], and it stimulates BK channels in human endothelial cells [34] through NOS activation. The NO donor also stimulates IBK of HCFs [25].
Signaling pathways involved in the effects of CO on BK currents of HCFs
CO is an endogenous modulator of the NO-cyclic GMP signaling system [52] and activates L-type calcium channels through NO- and cGMP dependent pathways [36]. Both CO [49] and NO [25] activate ion channels via the activation of sGC, which generates cGMP.
Our results demonstrated that 8-bromo-cGMP increased IBK (Fig. 3) and the CO-induced IBK stimulation effect in HCFs was blocked by the presence of a sGC blocker (ODQ) or a PKG blocker (KT5823). These results suggest that the stimulatory effects of CO are dependent on the sGC/cGMP/PKG signaling pathway. These results are consistent with that of a previous study on the effect of CO in human endothelial cells [34] and NO in HCFs [25].
CO is a weak stimulator of sGC compared with NO because CO binds to the sGC heme group with a lower affinity and can only weakly increase cyclic activity. The binding only results in a four- to six-fold activation of the enzyme. Unlike CO, NO increases the sGC activity 100–400-fold [37]. In previous reports, CO amplifies NO-induced cGMP levels seen with either CO or NO alone [53] and potentiates the elevation of NO-mediated cGMP [52]. Therefore, it seems that CO can function as a partial agonist to facilitate NO-mediated activation of sGC.
NO can exert many of its effects through cGMP-independent mechanisms: the c-AMP dependent pathways and S-nitrosylation. NO modulates BK channels through cAMP-dependent pathways in HCFs [25] and in rat cardiac fibroblasts [54]. NO also blocks Kv1.5 channels by S-nitrosylation [55]. Our results also demonstrated that the stimulating effects of CO of HCFs were mediated by cGMP-independent mechanisms; cAMP-dependent pathways and S-nitrosylation.
In our study, pre-treatment with a PKA blocker (KT5720) or an adenylate cyclase blocker (SQ22536) inhibited the effect of CO on the IBK in HCFs and cell membrane permeable cAMP, 8-bromo-cAMP treatment increased IBK (Fig. 4), which means that the cAMP-dependent pathway is also involved in the stimulating effect of CO on IBK in HCFs. These results are similar to the finding that NO increases IBK through PKG- and PKA-related pathways in HCFs [25] and cAMP-dependent vasodilators cross-activate the cGMP-dependent protein kinase pathway to stimulate BK channels in coronary artery smooth muscle cells [56].
We also found that CO could activate IBK in HCFs through S-nitrosylation, since a thiol-alkylating reagent, NEM, prevented CO stimulation effects on IBK and a reducing agent, DTT reversed the effect of CORMs on IBK (Fig. 5). This is the first report that an S-nitrosylation mechanism is involved in CO effects on IBK in a cardiac system and that a thiol residue could be the ultimate target of CO.
S-nitrosylation has emerged as an important and ubiquitous post-translational modification system, participating in cellular signaling (reviewed in Gonzalez et al.) [57]. Several reports exist on the effect of NO mediated through S-nitrosylation being implicated in all major functions of NO in the cardiovascular system [57-59]. Since S-nitrosylation signaling is involved in multiple physiological processes, it is expected that altered S-nitrosylation of specific ion channels may be relevant in some pathologic states, arrhythmia and heart failure.
Cardiac ion channels involved in excitation-contraction coupling are potentially regulated by S-nitrosylation [57]. In cardiac myocytes, S-nitrosylation is coupled to NOS activity for Nav1.5 channel activation [60]. NO inhibits L-type calcium channels by S-nitrosylation [61] and the α1-subunit of L-type calcium channels is constitutively S-nitrosylated in the mouse heart [62]. S-nitrosylation increases the slowly activating component of delayed rectifier K+ currents in a manner dependent on NOS [63,64] and Kv4.3 channels, which generates a transient outward K+ current [65].
Although BK channels are not expressed in the plasma membrane of cardiomyocytes, recent works showed that BK channels might localize at the sinoatrial node in the heart and contribute to the regulation of sinoatrial node cell automaticity. Application of paxilline significantly reduced the action potential firing of sinoatrial node cells and lengthened the diastolic depolarization phase of the action potential [66].
Considering fibroblast-myocyte electrotonic coupling [67], BK channels of HCFs and the CO effects on this the channel may lead to the discovery of novel therapeutic targets and the development of agents for improving outcomes of heart diseases.
In summary, the present study showed for the first time that CO stimulates BK channels of HCFs, which involves the activation of NO by NOS and the sGC/cGMP/PKG, adenylate cyclase/cAMP/PKA, and S-nitrosylation pathways.
 XML Download
XML Download