

 PDF
PDF Citation
Citation Print
Print


INTRODUCTION
Orofacial pain complaints are highly frequent and debilitating conditions concerning the neck, face, and head. These disorders are the responsibility of the clinician because the trigeminal nociceptive system is complex [1]. According to the gate-control theory proposed by Melzack and Wall [2] in 1965, the substaintia gelatinosa (SG) neurons or lamina II of the spinal cord are demonstrated as a fundamental section in regulation of nociceptive inputs from the periphery to the central nervous system (CNS). The trigeminal subnucleus caudalis (Vc), also referred to the medullary dorsal horn as a result of the considerable cytoarchitectural similarities with the spinal dorsal horn, is first-order relayed for the pain and temperature information from the dental and craniofacial regions [3,4]. It is generally accepted that the SG neurons of the Vc play a key role in receiving and modulating the painful stimuli from the orofacial pain through myelinated Aδ and unmyelinated C primary afferent fibers [5].
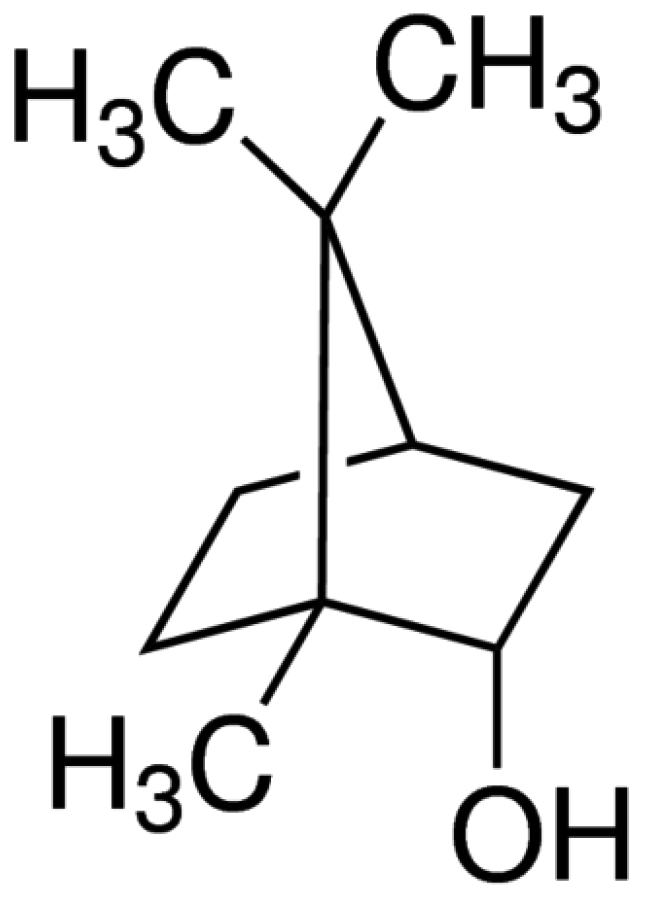
Borneol (L-borneol; C10H18O; Fig. 1), also known as Bing Pian and derived from several species of Artemisia and Dipterocarpaceae, is popularly used in food, fragrance, and folk medicine in Asian countries such as China, India, and Vietnam [6]. Borneol was approved by the US Food and Drug Administration (FDA) as a generally regarded as safe (GRAS) food flavoring [7]. With an acrid, bitter taste and cool properties, borneol is a fragrance ingredient in decorative cosmetics, shampoos, household cleaners, and detergents. Also, borneol has a broad range of traditional Chinese medical implications, particularly, in analgesia and antinociception [8]. In recent decades, this compound has gained the attention of neuroscientists in illuminating its empirical application based on scientific authority. It is reported that borneol alleviates mechanical hyperalgesia in the model of neuropathic pain [9], and attenuates oxaliplatin-induced neuropathic hyperalgesia [10]. This monoterpene was also evaluated for anti-nociceptive and anti-inflammatory activities in mice with the reduction of nociceptive behavior and inflammatory response [7]. Though borneol has been applied in traditional Chinese medicine for thousands of years and has also been the focus of attention in recent studies for its nociceptive effect [11,12], the relationship between borneol and orofacial pain treatment has not been completely reported yet. Thus, this study is focused on experiencing the actions of borneol on the SG neurons of the Vc and indicating relevant receptors using the whole-cell patch-clamp technique in mice.
METHODS
Animals and brain slice preparation
Experiments were performed under permission from the Institutional Animal Care and Use Committee of Jeonbuk National University (CBNU 2018-072, CBNU 2019-090). The experimental procedure has been described in detail in our previous studies [13]. After being decapitated, the brains of ICR mice were promptly exposed and drowned in ice-cold artificial cerebrospinal fluid (ACSF) comprising chemical compounds (in mM): 126 NaCl, 2.5 KCl, 2.4 CaCl2, 1.2 MgCl2, 11 D-(+)-glucose, 1.4 NaH2PO4, and 25 NaHCO3 (pH 7.3 to 7.4, bubbled with 95% O2 and 5% CO2). The brainstem was glued to an agar block and submerged in ice-cold ACSF. The coronal slices (180 to 220 μm in thickness) with the rostral part of Vc were excised by a vibratome (VT1200S; Leica Microsystems, Wetzlar, Germany). These brain slices were stored in the condition of oxygenated ACSF for at least one hour prior to electrophysiological recording.
Whole-cell patch-clamp recording
The coronal slices were delivered to the recording chamber ACSF submerged and constantly perfused at the rate of 4 to 5 ml/min. Visualization was performed from an upright microscope (BX51W1; Olympus, Tokyo, Japan) with differential interference contrast optics. The SG neurons of the Vc were discernible as a translucent band, just medial to the spinal trigeminal nucleus traveling across the lateral edge of the coronal slices.
Patch pipettes were prepared from the borosilicate glass capillary tubes (PG52151-4; WPI, Sarasota Bay, FL, USA) using a Flaming Brown puller (P-97; Sutter Instruments Co., Novato, CA, USA). High chloride pipette solution containing (in mM): 140 KCl, 1 CaCl2, 1 MgCl2, 10 HEPES, 4 Adenosine 5΄-triphosphate magnesium salt (Mg-ATP), and 10 EGTA (pH 7.3 with KOH) for boosting the chloride currents at the holding potential of –60 mV. Low chloride pipette solution containing (in mM): 136 C6H11KO7, 4 KCl, 1 CaCl2, 1 MgCl2, 10 HEPES, 4 Mg-ATP, and 10 EGTA (pH 7.3 with KOH) was applied for current-clamp experiment.
When charged by internal solution, the tip resistance of the patch pipette was approximately 4 to 7 MΩ. Once a gigaseal was formed between the pipette and the neuronal membrane, a slight negative pressure was applied to form the whole-cell mode. Signals were sequentially amplified by an Axopatch 200B (Molecular Devices, San Jose, CA, USA), digitized by an Axon Digidata 1550B (Molecular Devices). Acquired data were analyzed by the pClamp 10.6 software (Molecular Devices) and Origin 8 software (OriginLab Corp., Northampton, MA, USA).
Chemicals and statistics
Chemicals including borneol, picrotoxin, strychnine hydrochloride (strychnine), GABA, glycine, and ACSF compositions were purchased from Sigma-Aldrich (St. Louis, MO, USA), except for 6-cyano-7-nitro-quinoxaline-2,3-dione (CNQX), DL-2-amino-5-phosphonopentanoic acid (AP5), and tetrodotoxin citrate (TTX) by Tocris Bioscience (Avonmouth, Bristol, UK). Stocked drugs were dissolved in ACSF and tested at known concentrations.
In whole-cell voltage-clamp experiment, no less than 15 pA was considered to be responded by borneol to reduce possible error that may occur when measuring the currents. All data were presented as the mean ± standard error of the mean. A paired t-test was used to compare the mean values between the two groups. Statistical significance level was set as p < 0.05.
RESULTS
Seventy-three SG neurons from 34 mice (postnatal day 7 to 20) were experienced under the whole-cell patch-clamp technique including 67 neurons for voltage-clamp mode and 6 for current-clamp experiments.
Borneol induces repeatable inward currents on SG neurons
In whole-cell voltage-clamp experiment, 43 out of 54 SG neurons tested responded to 0.3 mM borneol and used for further analysis.
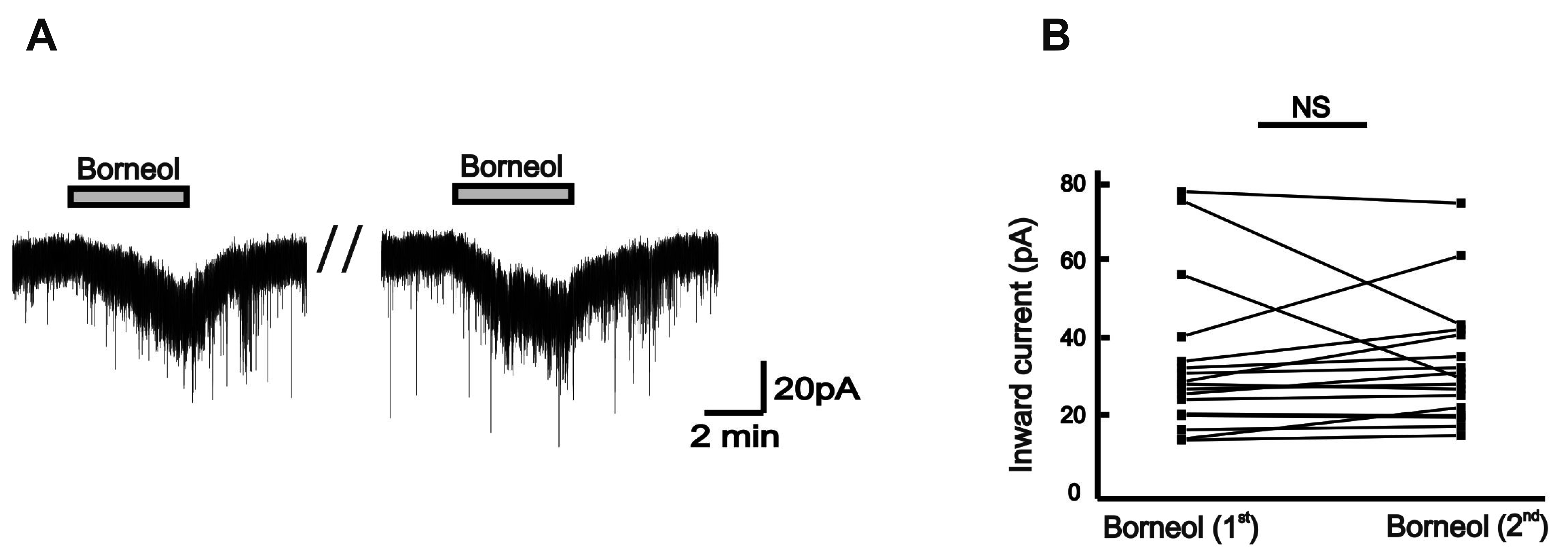
Under the high chloride pipette solution, borneol was successively introduced into the bath to examine how SG neurons of the Vc respond to borneol. No significant difference was found in the borneol-induced inward currents between the first and second applications (Fig. 2A). In 17 neurons tested, the mean amplitude of inward current (34.8 ± 4.71 pA) induced by the first application of borneol was similar to that of the second application (34.8 ± 3.85 pA, n = 17, p > 0.05; Fig. 2B) indicating that SG neurons of the Vc are not desensitized by the consecutive applications of borneol.
Borneol directly acts on SG neurons
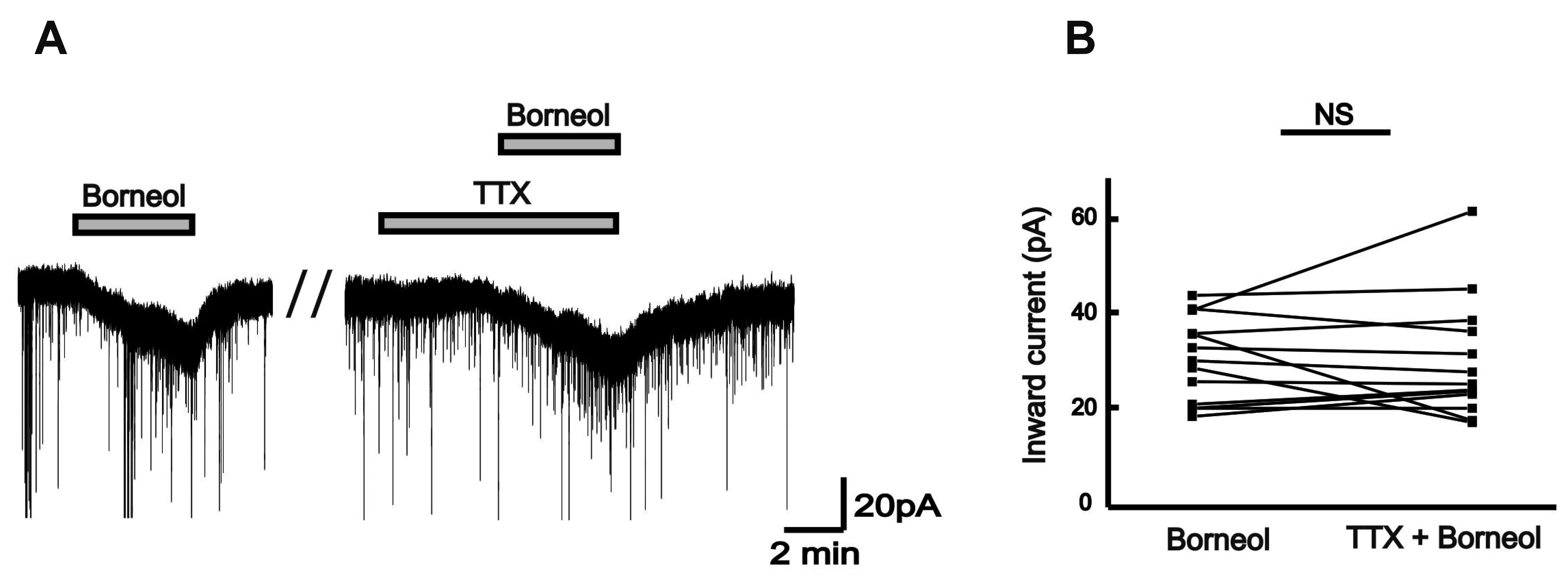
TTX, a potent selective blocker of sodium channel mediated by the membrane potential, can interfere with the generation of action potentials and depress the propagation of impulses in excitable membranes [14]. TTX acts on neither the release of the transmitter from nerve terminals nor the sensitivity of the postsynaptic membrane to the transmitter. Therefore, TTX applications are extremely useful for studies of the mechanism of transmitter release and of the mechanism whereby the transmitter interacts with the postsynaptic membrane [15]. In this study, TTX was co-applied with borneol to examine whether borneol acts on SG neurons via action potentials generated. The borneol-induced inward currents were conserved in the presence of 0.5 µM TTX in 13 SG neurons (Fig. 3A). There was a similarity in the mean amplitude of borneol-induced inward current between the absence (31.4 ± 2.47 pA) and the presence (31.2 ± 3.54 pA, n = 13, p > 0.05; Fig. 3B) of TTX. This result indicates that borneol directly acts on the postsynaptic SG neurons.
Borneol activates GABAA receptor and/or glycine receptor
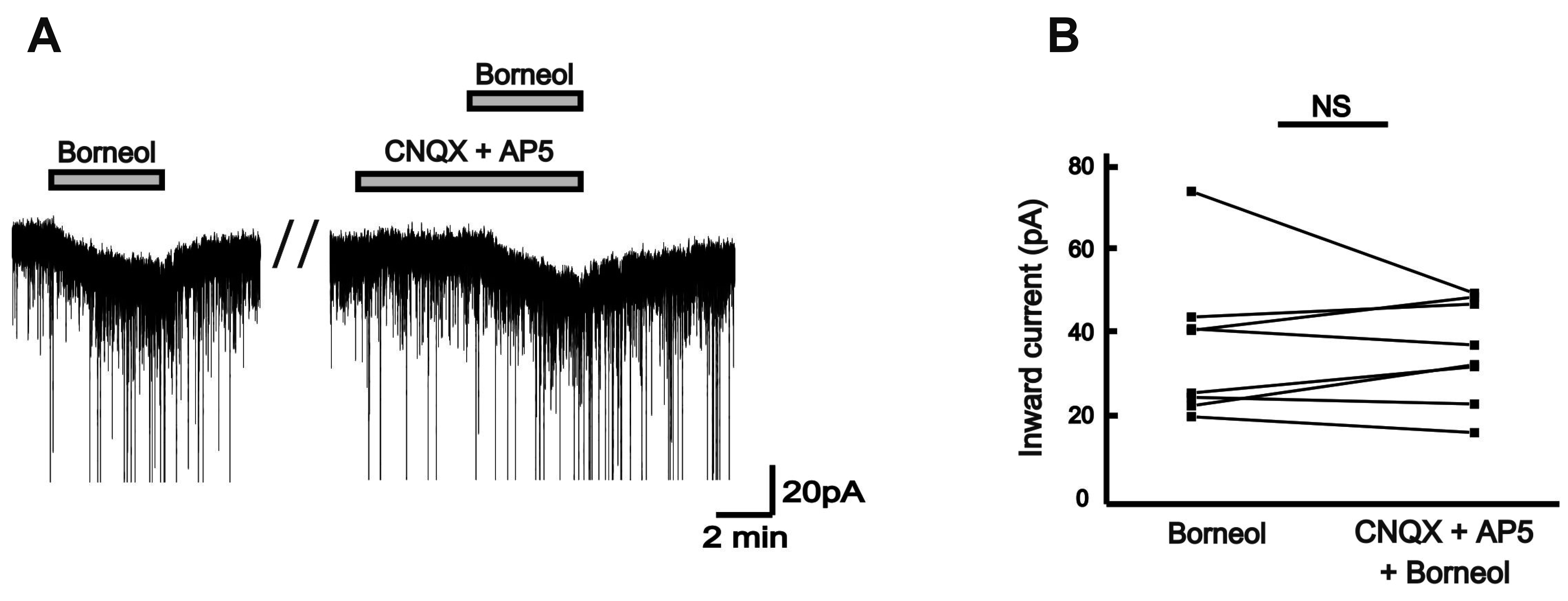
To consider the action of borneol is mediated via ionotropic glutamate receptors, borneol was examined in a cocktail of CNQX, a non-NMDA glutamate receptor antagonist and AP5, an NMDA receptor antagonist. The borneol-induced inward currents were not affected by the presence of CNQX and AP5 in 8 tested neurons (Fig. 4A). The mean inward current (37.7 ± 6.33 pA) by borneol was proximately similar to that in the presence of AP5 (20 µM) and 10 µM CNQX (36.9 ± 4.36 pA, n = 8, p > 0.05; Fig. 4B).
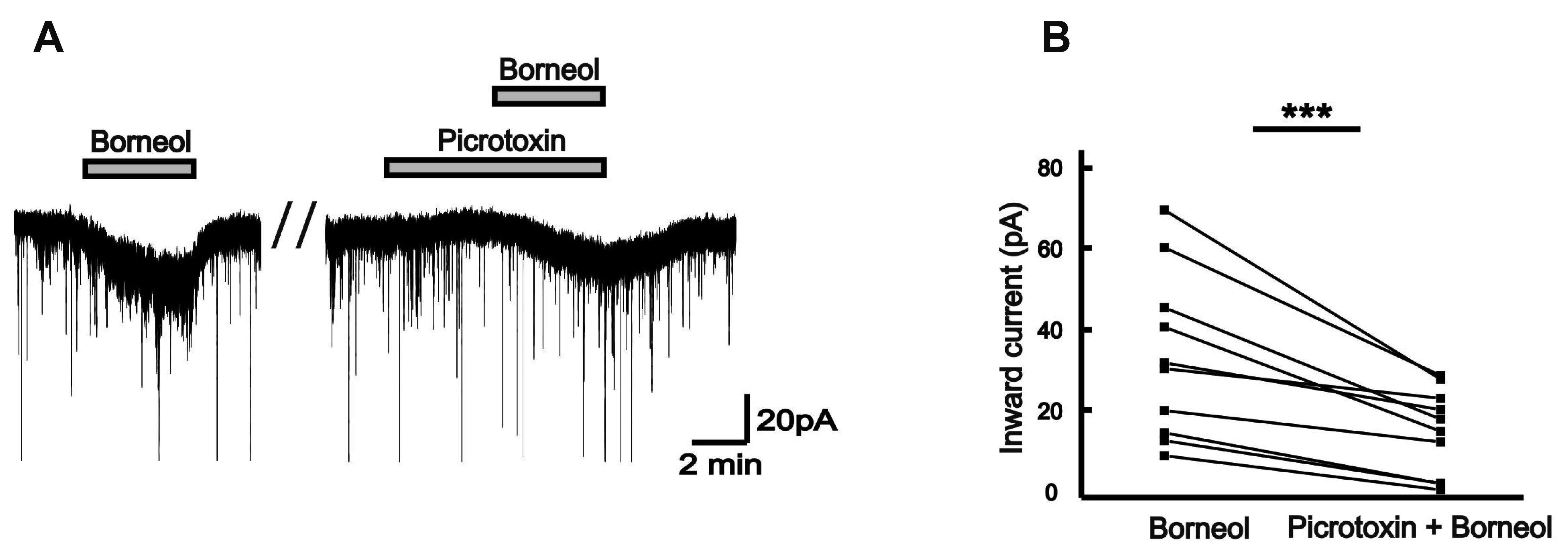
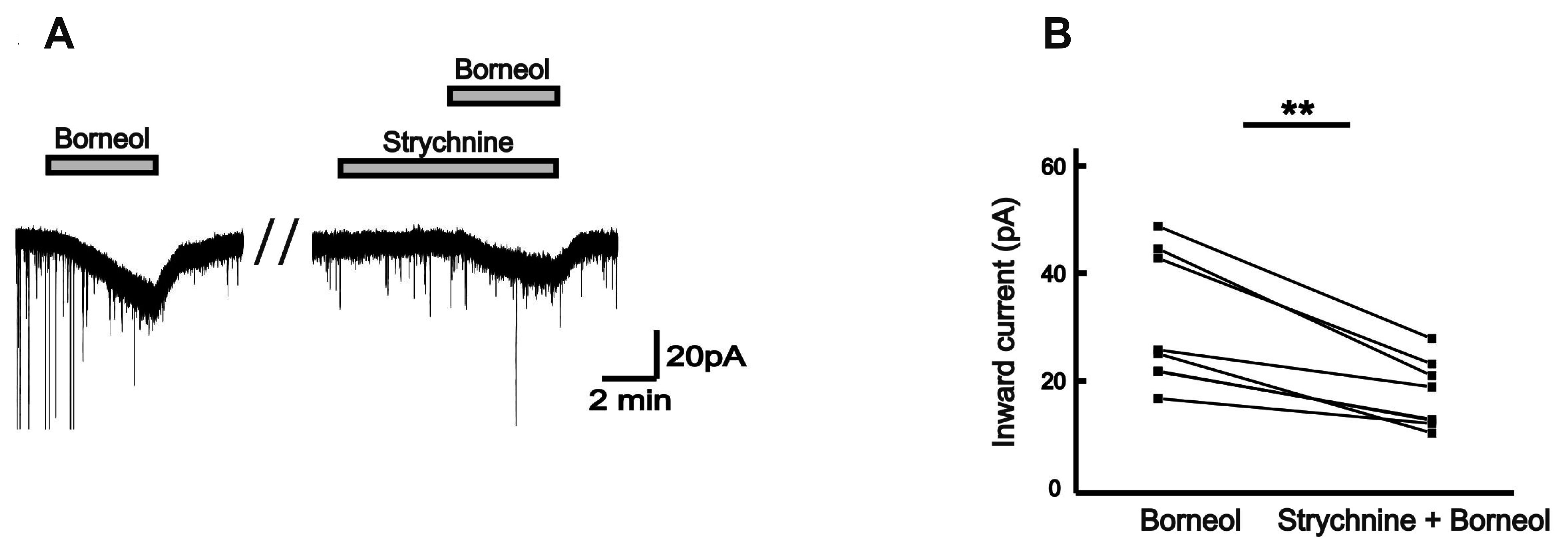
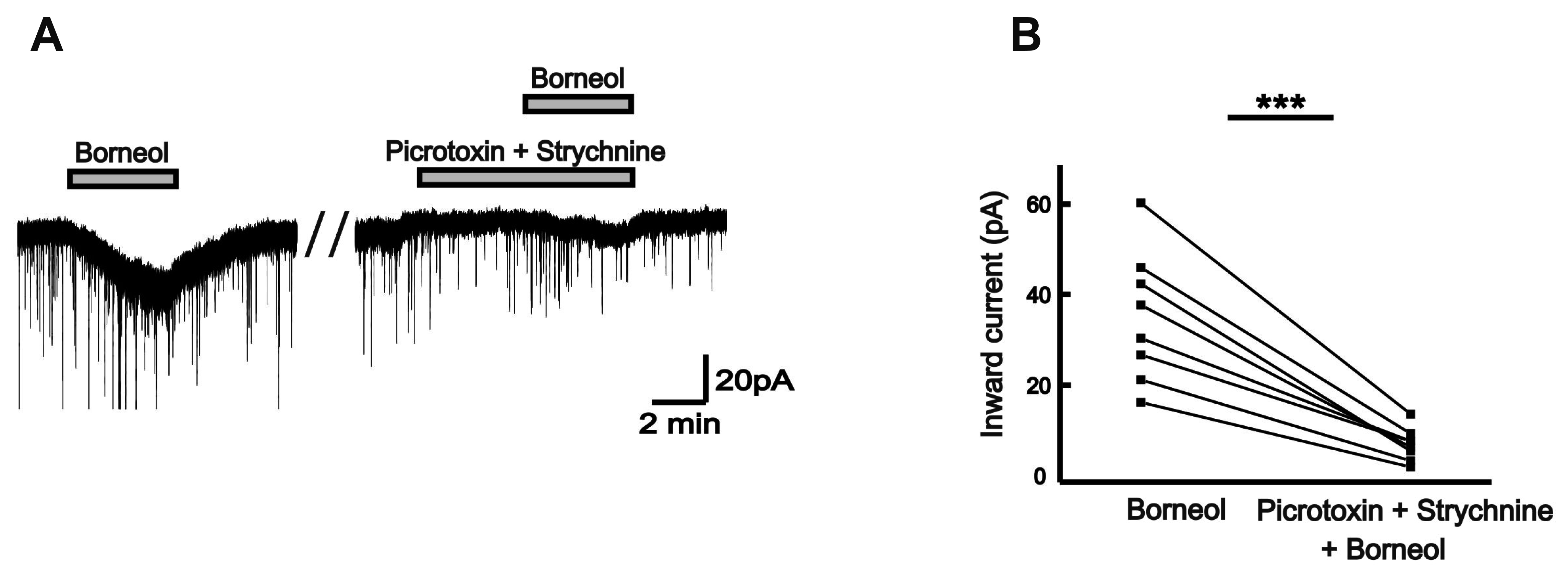
Picrotoxin, a non-competitive GABAA receptors antagonist, is often used as a tool to prove the behavior of GABA receptors [16]. In the presence of 50 µM picrotoxin, the borneol-induced currents tended to reduce, compared with those in the absence of picrotoxin in the same neurons (Fig. 5A). The mean inward currents induced by borneol were 37.6 ± 6.49 pA and 18.0 ± 3.27 pA in the absence and the presence of picrotoxin, respectively (n = 10, p < 0.001; Fig. 5B). Additionally, a competitive antagonist of glycine receptor, strychnine was applied for testing the relationship between the borneol response and the strychnine-sensitive glycine receptors [17]. The borneol-induced currents experienced significant decrease when 2 µM strychnine was applied prior to the borneol (Fig. 6A). The mean inward current (31.9 ± 4.40 pA) by borneol was reduced in the presence of strychnine (18.3 ± 2.24 pA, n = 7, p < 0.01; Fig. 6B). Also, the borneol-induced inward currents were almost blocked by co-application of picrotoxin and strychnine (Fig. 7A). The mean inward current (36.5 ± 5.10 pA) induced by borneol was almost depressed to 8.31 ± 1.28 pA in the presence of picrotoxin and strychnine (n = 8, p < 0.001; Fig. 7B). These results indicate that borneol-induced actions are mediated by activation of GABAA and/or glycine receptors but not by ionotropic glutamate receptors.
Borneol enhances GABA-mediated actions
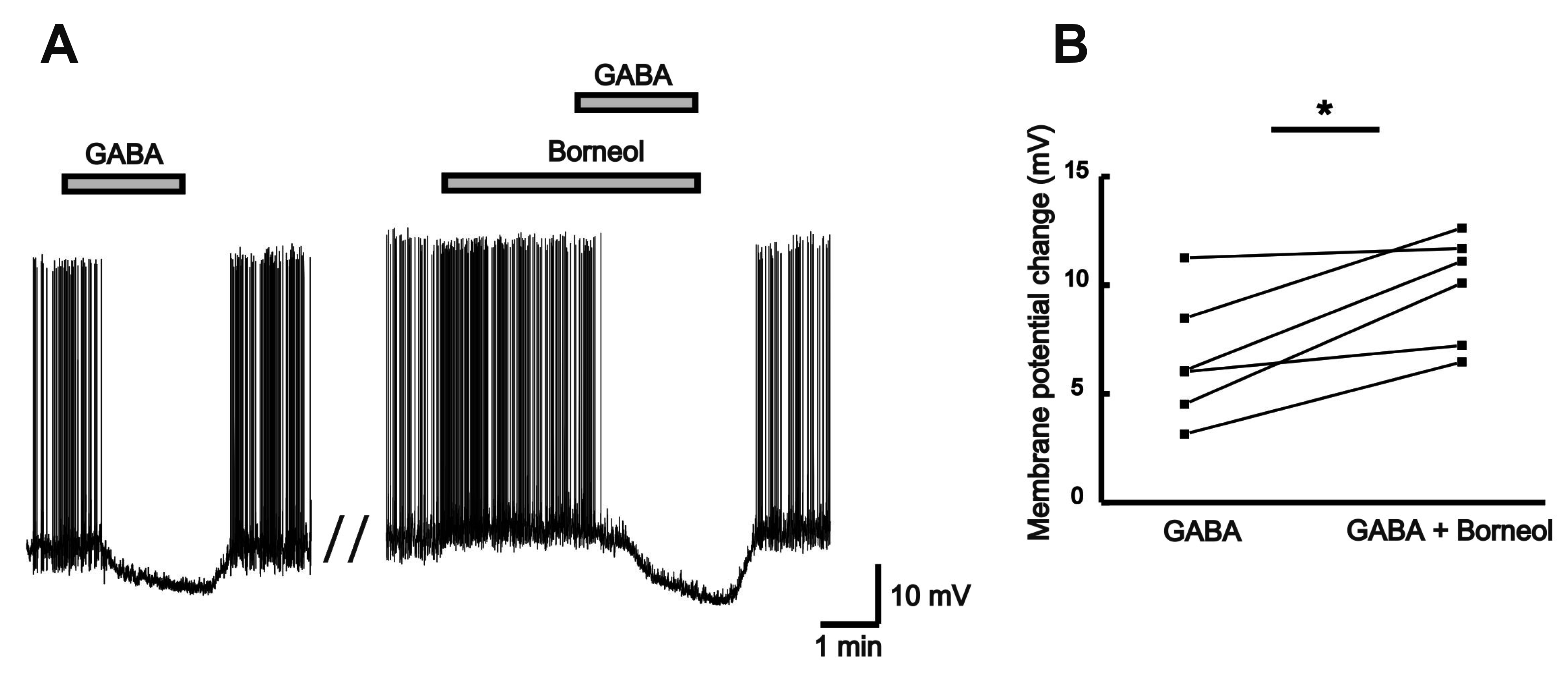
Although controversy persisted well into the function of GABA and glycine, it is now widely admitted that GABA and glycine are the most crucial inhibitory neurotransmitters in the CNS [18]. Under the high chloride intracellular condition, GABA and glycine can induce inward currents as a result of the chloride ion out-flux. Lower concentration of borneol (0.2 mM) not producing detectable inward currents was co-applied to check the effect of borneol on the GABA or glycine responses. Fig. 8A exhibits a representative trace showing a potentiation effect between GABA (30 µM) and borneol (0.2 mM). The mean GABA-induced inward current (164 ± 40.8 pA) was increased in the presence of borneol (300 ± 64.9 pA, n = 5, p < 0.05; Fig. 8B). Fig. 8C exhibits a representative trace showing no borneol effect on glycine-induced response. The mean inward currents (132 ± 33.5 pA) induced by glycine (30 µM) was not changed in the presence of borneol (0.2 mM, 130 ± 30.5 pA, n = 8, p > 0.05; Fig. 8D).
To confirm whether borneol affects the GABA-induced hyperpolarization, borneol was applied in current-clamp mode under low chloride pipette solution. GABA (30 µM) induced hyperpolarization in all neurons tested and the GABA-induced hyperpolarization was enhanced by borneol (Fig. 9A). The mean hyperpolarization by GABA (6.58 ± 1.19 mV) was increased in the presence of borneol (0.3 mM, 9.87 ± 1.02 mV, n = 6, p < 0.05; Fig. 9B).
DISCUSSION
Giving a brief summary, borneol-induced non-desensitizing inward currents were preserved in the presence of TTX or CNQX and AP5, but suppressed by the presence of strychnine, a glycine antagonist and picrotoxin, a GABAA antagonist. Additionally, borneol enhanced the GABA-mediated inward currents under the high chloride pipette solution and the GABA-induced hyperpolarization in the low chloride internal solution on the SG neurons of the Vc in mice.
In this study, we used 0.3 mM borneol. It has been reported that the half-maximal effective concentration (EC50) of borneol to enhance GABA effect in Xenopus laevis oocytes and half maximal inhibitory concentration (IC50) of borneol to inhibit human TRPA1 mediated cationic currents in heterologous expression systems like Xenopus oocytes or in neurons cultured from trigeminal ganglia were 248 µM and 200 µM, respectively [19,20].
Although borneol showed GABA or glycine mimetic actions, the responses were shown much slower than that of typical GABA or glycine. In some whole-cell patch-clamp studies, borneol-mediated responses were evaluated around 3 to 5 min after borneol application on the members of transient receptor potential family such as TRPA1 and TRPM8 [20,21]. So, further studies might be needed to confirm the precise mechanism how borneol activates GABA and glycine receptors.
Borneol, a bicyclic organic compound and a terpene derivative, is historically termed as borneocamphor. In traditional Chinese medicine, borneol is classified as “Herbs that Open Orifices” and is commonly used to treat conditions associated with nociception [22]. However, traditional herbal remedies have encountered significant skepticism because these treatments are mostly based on anecdotes and clinical observations instead of verifiable scientific trials. In recent decades, scientists have shown increasing interest in deciphering the nociceptive mechanism responsible for the borneol clinical effect. Borneol has demonstrated its analgesic and anti-inflammatory effects on the photodynamic therapy of acne [23]. A study measuring nociception by acetic acid, formalin, hot plate, and grip strength tests has proved that borneol produces a significant reduction of nociceptive behavior and proposes this terpene as a substantial central and peripheral antinociceptive agent [7]. Borneol can inhibit TRPA1, a member of TRP family, as a key player in neuropathic pain and inflammation [12]. Also, borneol has demonstrated a considerable antihyperalgesic activity in the mouse model of chemotherapy-induced neuropathic pain [10]. This body of evidence somewhat demonstrates that borneol has been a traditional pain management treatment for thousands of years.
The operation of neuronal networks depends on the balance of the excitatory and inhibitory transmission, in which GABA and glycine function as the main inhibitory neurotransmitters in the spinal cord and dorsal horn. There is an array of natural terpene compounds of plant origin, including citral, eucalyptol, or linalool proven to influence the function of ionotropic GABAA receptors [13]. Glycine largely modulates the fast inhibitory neurotransmission in the spinal cord and brainstem through specific glycine receptors popular in the postsynaptic membrane [17]. GABAA or glycine receptors are a ligand gated chloride channel, the opening of which results in the membrane hyperpolarization, and depression of firing action potentials [17,24].
As the main inhibitory neurotransmitters in CNS, it is widely believed that GABA and glycine have a major role in controlling nociceptive processing. A study concerning fear recall and anxiety-like behaviors indicates that the currents evoked by GABA are enhanced by borneol in mice hippocampal slices and the enhancement is reversed in the presence of bicuculline, a GABAA receptors antagonist [25]. Borneol has also been proven to ameliorate mechanical hyperalgesia in models of chronic inflammatory and neuropathic pain via increasing GABAergic transmission in the spinal cord [9]. A study on the content changes of excitatory and inhibitory neurotransmitters in corpus striatum of rats using single-applied of borneol or combining it with diazepam indicates that borneol significantly increased the content of glycine and GABA but not aspartate and glutamate [26]. From our results, borneol is demonstrated to activate GABAA receptor and/or glycine and possess a potentiation effect with GABA but not glycine. This might be explained that GABA receptors are more sensitive than glycine receptors to positive allosteric modulation by borneol. This mechanism is also indicated in an experiment on Xenopus oocyte revealing that borneol showed an inhibitory neurotransmission enhancement via a stronger potentiation effect with GABA than that with glycine [27].
The majority of SG neurons are inhibitory including GABA and glycine, in that approximately one third of SG neurons are GABAergic [28]. The Vc contributes substantially in the transmission of non-discriminative touch, thermal sensations and nociception from the face and oral cavity [3,4]. For these reasons, the SG neurons of the Vc is considered as a principal region for regulating the orofacial nociceptive information via unmyelinated C and myelinated Aδ fibers. Knowledge about the response of SG neurons of the Vc to traditional Chinese medicine-based on agents may set scientific documentation as well as develop a novel approach for understanding their effect on the orofacial nociception.
Together, we demonstrate that borneol can directly activate GABAA receptor or glycine receptor and enhance GABA-mediated actions on the SG neurons of the Vc in mice, and suggest that borneol may serve an agent concerning to inhibitory synaptic transmission of the CNS and be a therapeutic target for the orofacial pain treatment.
 XML Download
XML Download