

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
The transient receptor potential (TRP) channels are known to play a significant role in intracellular calcium regulation. We have previously reported that among TRP families, the TRP canonical (TRPC) 5 channel is crucial not only in calcium regulation, but also extracellular pH sensing [1]. We have identified that the G-protein coupled receptors (GPCR) pathway induces TRPC5 activation independent from the activation by the trivalent cations, La3+ and Gd3+ [23]. The potentiation of TRPC5 is reported to be enhanced by several different factors, such as extracellular phospholipids [4], heavy metals [5], oxidized glutathione [6], and nitric oxide [7], and desensitized by protein kinase C-mediated phosphorylation [8].
Englerin A (EA) is a novel natural product that selectively inhibits the growth of renal cell carcinoma cell lines [9]. Recently, it has been used as an activator of TRPC4 and TRPC5 channels. A recent study reported that G-protein signaling does not take part in the channel activation by EA [9]. The study also uncovered that the application of EA on TRPC4 or 5-overexpressed human embryonic kidney (HEK) cells activates the channels and induces cell death in consequence of increasing intracellular calcium levels [9]. Such results were supported by our previous studies where we reported Gαi, oxidized glutathione and constitutively active mutant of TRPC4, 5 increased intracellular Ca2+ [3610] and used TRPC4-GCaMP6 to find that EA increased calcium levels near TRPC4 channels [11]. However, the mechanism of EA-induced activation of the ion channels is unclear. A report suggests that the phenyl ring in EA plays a significant role in the activation of the L-type calcium channel [12]. In the present study, we have identified the EA-interaction sites in TRPC5 using a patch clamp technique by which we recorded the EA-induced current increases of TRPC5 mutants. The present study provides a plausible and novel mechanism of TRPC5 channel activation.
METHODS
Cell culture and transient transfection
Human embryonic kidney (HEK293) cells (ATCC, Manassas, VA, USA) were maintained according to the supplier's recommendations. For transient transfection, cells were seeded in 12-well plates. The following day, 0.5 µg/well of pcDNA vector containing the cDNA for TRPC5-EGFP was transfected into the cells using the transfection reagent FuGENE 6 (Promega, Madison, WI, USA) according to the manufacturer's protocol. After 30 to 40 h, the cells were trypsinized and used for whole-cell recording.
Molecular biology
Plasmid containing human TRPC5 gene was given kindly by Dr. Shuji Kaneko in Japan. Point mutations in TRPC5 were introduced using the QuickChange site-directed mutagenesis kit (Agilent Technologies, Santa Clara, CA, USA) and appropriate primer sets. Sequences of the mutants were confirmed by DNA sequencing.
Electrophysiology
The whole-cell patch clamp was performed to measure the TRPC channel current in HEK cells. The transfected cells were trypsinized from the culture well and attached to coverslips in the small chamber on an inverted microscope for 10 min prior to patch recording. The currents were recorded using an Axopatch 200B patch clamp amplifier (Molecular Devices, San Jose, CA, USA). The bath solutions were constantly perfused with normal Tyrode (NT) solution at a rate of 1 to 2 ml/min. Glass microelectrodes with 2 to 4 MΩ resistance when filled with internal solution were used to obtain gigaohm seals. For some experiments, after establishing the whole-cell configuration, the external solution was changed from NT to Cs+-rich solution for recording TRPC5 current in HEK293 cells. The extracellular ion composition was altered to examine the activity of TRPC5 channel, especially for cations with high permeability. Voltage ramp pulse was applied from +100 to −100 mV for 500 msec at −60-mV holding potential at every 20 sec. Experiments were performed at room temperature (20℃–24℃). The pCLAMP (version 10.2) and Digidata 1440A (Axon Instruments) software programs were used for data acquisition and the application of command pulses. The data were filtered at 5 kHz and displayed on a computer monitor. The data were analyzed using pCLAMP (version 10.2) and OriginPro 8 (OriginLab, Northampton, MA, USA).
Western blotting
Cells were plated in 6-well dishes. Lysates were prepared in lysis buffer (0.5% Triton X-100, 150 mM NaCl, 50 mM HEPES, 2 mM MgCl2, 2 mM ethylenediaminetetraacetic acid [EDTA], pH 7.4) via passage 15 to 20 times through a 26-gauge needle. After lysates were centrifuged at 13,000 ×g for 10 min at 4℃, the protein concentration in the supernatants was determined. The extracted proteins were separated on sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to nitrocellulose membranes. The following primary antibodies were used for Western analysis: rabbit antibody against GFP (A-11122; Invitrogen, Carlsbad, CA, USA) and mouse antibody against β-tubulin (T-4026; Sigma Aldrich, St. Louis, MO, USA). Image-J Software was used to quantify the level of protein expression.
Western blotting and surface biotinylation
Cells were plated in 6-well dishes. Lysates were prepared in lysis buffer (0.5% Triton X-100, 150 mM NaCl, 50 mM HEPES, 2 mM MgCl2, 2 mM EDTA, pH 7.4) via passage 15 to 20 times through a 26-gauge needle. After lysates were centrifuged at 13,000 ×g for 10 min at 4℃, the protein concentration in the supernatants was determined. The extracted proteins were separated on SDS-PAGE and transferred to nitrocellulose membranes. The following primary antibodies were used for Western analysis: rabbit antibody against GFP (A11122; Life Technologies) and mouse antibody against β-tubulin (T-4026; Sigma Aldrich). Image-J Software was used to quantify the level of protein expression.
For surface biotinylation, HEK293 cells were transfected with wild-type (WT) TRPC5 or mutants. After 24 h, cell membrane protein biotinylation was performed. Cells were washed in phosphate-buffered saline (PBS) and then incubated with 0.5 mg/ml Sulfo-NHS-LC-Biotin in PBS on ice for 30 min. Free biotin was quenched by 100 mM glycine in PBS, followed by lysing procedure at 4℃ for 30 min. Lysates were centrifuged at 13,000 ×g for 10 min at 4℃, and protein concentration in the supernatants was determined. A 50% of Pierce™ Avidin beads (Thermo Scientific, Waltham, MA, USA) was added to cell lysates. After incubation for 1 h at room temperature, beads with protein were washed 3 times with 0.5% Triton X-100 in PBS, and proteins were extracted in sample buffer. Collected proteins were then analyzed by Western blot.
Solutions and drugs
For recordings of the TRPC channels, we used NT solution and Cs+-rich solution. The NT solution contained 135 mM NaCl, 5 mM KCl, 2 mM CaCl2, 1 mM MgCl2, 10 mM glucose, and 10 mM HEPES with pH of 7.4 adjusted with NaOH. The Cs+-rich external solution contained equimolar CsCl instead of NaCl and KCl with pH adjusted to 7.4 using CsOH. The internal solution contained 140 mM CsCl, 10 mM HEPES, 0.2 mM Tris-GTP (Tris-guanosine 5′-triphosphate), 0.5 mM EGTA, and 3 mM Mg-ATP (adenosine 5′-triphosphate) with pH of 7.3 adjusted with CsOH and used 0.2 mM GTPγS (guanosine 5′-triphosphate-γ). All reagents were purchased from Sigma Aldrich).
Statistics
All statistical analysis was done with GraphPad Prism 5 (GraphPad Software Inc., San Diego, CA, USA). Results are presented as means ± standard error of mean. Statistical data were obtained by paired or unpaired Student's t-test and ANOVA. The p values less than 0.05 were considered statistically significant. The number of whole cell recordings is indicated by n.
RESULTS
Identification of putative EA-interaction sites in TRPC5 ion channel
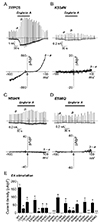
To investigate on the mechanism of TRPC5 ion channel activation by EA, we first generated TRPC5 mutants in which selected amino acids within the pore region are mutated. The mutants were generated according to their propensity to sense H+ or OH− ions. The current was recorded under the condition of 150 mM of external Na+ and without GTPγS in the patch pipette, which is an activator of TRPC5. Each mutant was over-expressed in HEK293 cells and its whole-cell currents were recorded under the same condition. When 100 nM of EA was applied, the current change (b−a) of TRPC5WT was −161.63 ± 19.50 pA/pF (n = 6) at −60 mV (Fig. 1A, E). As shown in Fig. 1E, the mutants showed their varying responses to EA, and their response patterns could be divided into three categories (Fig. 1E). First, there was no current response to EA in six mutants: TRPC5Y542F, TRPC5C553A, TRPC5K554N, TRPC5C558A, TRPC5H594N, and TRPC5E598Q. We previously reported that we could not observe whole-cell TRPC5 currents in TRPC5Y542F, TRPC5C553A, and TRPC5C558A overexpressed in HEK293 cells [13]. Western blot analysis also showed remarkably reduced expression of total TRPC5Y542F protein and expressed protein at the plasma membrane (Supplementary Fig. 1), and TRPC5C553A and TRPC5C558A protein at the plasma membrane [2]. The mutants TRPC5K554N (Fig. 1B), TRPC5H594N (Fig. 1C), and TRPC5E598Q (Fig. 1D) were expressed in HEK293 cells. Compared to WT, the mutant H594N and E598Q showed similar total protein and expressed protein in the plasma membrane whereas K554N was decreased at both total and expressed protein at the plasma membrane (Supplementary Fig. 1). Their EA-induced currents, however, were increased only to −0.30 ± 0.42 pA/pF (n = 5), −1.42 ± 0.35 pA/pF (n = 5), −0.10 ± 0.63 pA/pF (n = 5), respectively. The positive effect of EA on TRPC5 was abolished in these three mutants. The mutants in the second group showed that the current changes in the middle range from −10 to −30 pA/pF. The EA-induced currents were not abolished, but reduced (Fig. 1E). The mutant TRPC5(R545N) showed relatively substantial reduction in EA-induced current increases, yielding −10.08 ± 6.17 pA/pF (n = 5) (panel a in Fig. 2A). Likewise, TRPC5Y539F (−27.26 ± 5.04 pA/pF [n = 5]), TRPC5D548N (−28.14 ± 8.31 pA/pF [n = 5]), TRPC5E549Q (−30.80 ± 16.25 pA/pF [n = 5]; panel b in Fig. 2A), TRPC5K560N (−17.24 ± 3.64 pA/pF [n = 5]; panel c in Fig. 2A), and TRPC5E570Q (−13.69 ± 3.97 pA/pF [n = 5]; panel d in Fig. 2A) showed similar reductions in EA-induced current increases. Unlike the mutants in the second group, the EA-induced current increases of the third group, which are TRPC5Y541F (−68.25 ± 17.65 pA/pF [n = 5]), TRPC5E543Q (−81.45 ± 12.43 pA/pF [n = 5]; panel a in Fig. 2B), TRPC5R557N (−97.60 ± 8.21 pA/pF [n = 5]; panel b in Fig. 2B), TRPC5E559Q (−60.19 ± 17.64 pA/pF [n = 5]), TRPC5Y586F (−81.45 ± 12.43 pA/pF [n = 5]), TRPC5R593N (−68.86 ± 13.28 pA/pF [n = 5]; panel c in Fig. 2B), and TRPC5E595Q (−62.98 ± 6.87 pA/pF [n = 5]; panel d in Fig. 2B) were reduced to greater amplitudes. These results suggest that the three specific residues, Lys-554, His-594, and Glu-598, are putative EA interaction sites in the TRPC5 ion channel. On the other hand, it is difficult to rule out the possibility that EA has no effect on the three mutants because there are permeability changes or modulation of gating due to mutation.
Functional intactness of TRPC5 pore mutants
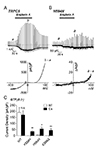
To examine whether TRPC5K554N, TRPC5H594N, and TRPC5E598Q mutants are functionally intact, we used GTPγS-included internal solution, while other conditions were maintained. Application of EA on the TRPC5WT-expressing HEK293 cell increased the current by −190.67 ± 39.88 pA/pF (n = 6) (Fig. 3A, C). The EA-induced current increases of TRPC5K554N, TRPC5H594N, and TRPC5E598Q were −0.32 ± 0.39 pA/pF (n = 5), −5.47 ± 0.61 pA/pF (n = 5), and −0.55 ± 0.51 pA/pF (n = 5), respectively (Fig. 3B, C). Compared to TRPC5WT, the three mutants lost channel activation by EA even with GTPγS, which is a TRPC5 activator (Fig. 3C).
Since TRPC5 has higher permeability to Cs+, we used Cs+-rich external solution in addition to GTPγS in the internal solution. The EA-induced current increases of TRPC5WT was −170.61 ± 35.35 pA/pF (n = 6). With Cs+-rich solution, the current increase of the three mutants were restored, although one-fourth of WT. Each mutant, TRPC5K554N, TRPC5H594N, and TRPC5E598Q showed current increases by −34.66 ± 8.42 pA/pF (n = 5), −55.96 ± 12.96 pA/pF (n = 5), and −44.64 ± 12.50 pA/pF (n = 5), respectively (Fig. 3C, Supplementary Fig. 1). These results suggest that three mutants did not lose channel function completely.
Since muscarinic stimulation activates TRPC5, we applied CCh without any expression of muscarinic receptor prior to coexpression with the receptor. Whole-cell currents from each mutant showed the typical double rectifying I/V curve although the amplitude was less than 2 pA/pF (Fig. 4A) except for TRPC5H594N. Next, we co-expressed M3 muscarinic receptor with each mutant and applied CCh to activate TRPC5 channels (Fig. 4B). Whole-cell currents from each mutant showed the typical double rectifying I/V curve except for TRPC5H594N. Finally, CCh was applied under the condition of 140 mM of external Cs+ to increase the TRPC5 currents. Since TRPC5 has higher permeability to Cs+, changing NT to Cs solution increased TRPC5 currents. CCh application further increased TRPC5 current. Cs+-induced basal current (Fig. 4C) and CCh-induced current (Fig. 4D) at each mutant were smaller than that of WT TRPC5. These results suggest that each mutant has an electrophysiological function, although the mutation affected the pore region and decreased the level of their response to TRPC5 agonists.
Activation of TRPC5 pore mutant E595Q/E598Q
Since TRPC5 E595Q/E598Q revealed a significant receptor-activated current [14], we generated E595Q/E598Q double mutant and tested the effect of CCh instead of histamine. Without M3 receptor expression, CCh induced similar currents in E595Q/E598Q double mutant compared with WT (Supplementary Fig. 2). Cs+-induced basal current was smaller in E595Q/E598Q double mutant (Supplementary Fig. 2B). Under the condition of Cs+, CCh also induced smaller currents compared with WT (Supplementary Fig. 2C). We investigated whether E595Q/E598Q double mutant responds to EA (Supplementary Fig. 3). E595Q/E598Q double mutant has a similar response to EA as WT (Supplementary Fig. 3). The I/V curve showed the typical double rectifying shape (panel b in Supplementary Fig. 3A). These results suggest that the response to EA might be different from the response to other agonists, such as Cs+ or muscarinic agonists. In addition, the permeability change does not explain the response of three mutants to EA.
Activation of TRPC5 pore mutants by higher concentration of EA
Since the response to EA might be different from the response to other agonists, such as Cs+ or muscarinic agonists, we tested whether the sensitivity to EA changed due to mutations. When 1 µM EA was applied to HEK293 cells expressing WT TRPC5, the cells did not survive such concentrated treatment. When 1 µM of EA was applied to HEK cells expressing mutant TRPC5, however, the current increased, and the current amplitude was similar to that of WT TRPC5 stimulated by 100 nM of EA (Fig. 5). These results suggest that the response to EA might be different from the response to other agonists, such as Cs+ or muscarinic agonists, and the sensitivity to EA decreased in TRPC5K554N, TRPC5H594N, and TRPC5E598Q mutant, at least ten-fold.
DISCUSSION
The TRPC5 has been known to be a receptor-activated cationselective and [Ca2+]i-dependent channel [1315]. The channel is activated at 100 nM or higher intracellular calcium concentration, but does not depend on the calcium store depletion [1315]. In addition, there is a growing evidence that TRPC5 channel is also activated by EA. In the present study, we have identified the EA-interaction sites within the pore region of TRPC5, along with other sites that are likely to play an ancillary role in EA-mediated channel regulation. While Lys-554, His-594, and Glu-598 might be likely to be direct interaction sites, Arg-545, Glu-549, Lys-560, Glu-570, Arg-593, and Glu-595 are likely to play indirect roles. When we tested whether the sensitivity to EA changed due to mutations, 1 µM of EA increased the current amplitude similar to that of WT TRPC5 stimulated by 100 nM of EA (Fig. 5). The sensitivity to EA decreased in TRPC5K554N, TRPC5H594N, and TRPC5E598Q mutant at least ten-fold, contrary to our expectation that the response to EA would be completely lost. Thus, we could not rule out that mutations just alter channel conformation. In case of K554N mutant, even though the expression level of total protein is low as in Y542F, Cs+ induced currents under the condition of GTPγS activator. A similar tendency was obtained in K554R mutant. Further studies are needed to determinew the role of K554 in the TRPC5 activation of EA using more mutants.
TRPC5 has 65% sequence identity to TRPC4 but only 45% sequence identity to TRPC1 [1617]. In this study, we observed that while TRPC5 was activated by EA, cell expressing TRPC1 did not respond to EA (Supplementary Fig. 4). We also showed that TRPC4 was activated by EA [11]. In addition, EA activated TRPC1/4 and TRPC1/5 heteromer and induced cell toxicity via Na+ influx instead of Ca2+ influx [1819]. As shown in Supplementary Fig. 4, the three residues in TRPC5 critical for interaction with EA are found different in TRPC1. This provides credible evidence of the different responses of the TRPC1 channel to EA application. The fact that EA activated TRPC1/4 and TRPC1/5 heteromers suggest that activation by EA does not require four protomers of TRPC4 and TRPC5. The recent report uncovers the mechanism of EA-mediated L-type calcium channel activation [12]. EA is reported to have selective binding affinity for dihydropyridine in Cav1.2 channels and modulate calcium signaling in muscle cells and vascular tissue [12]. Although we simply utilized the charged amino acids in the current study, a recent report suggests that EA not only acts upon OH− or O but also phenyl ring inducing conformational change in L-type calcium channel activation [12]. Three tyrosine mutants, TRPC5Y539F TRPC5Y541F, and TRPC5Y586F, responded to EA to a lesser degree compared with TRPC5WT. To our disappointment, we could not test whether TRPC5Y542F mutant responds or not because TRPC5Y542F mutant did not show the typical double rectifying I/V curve to any agonist, and the expression level of total protein was significantly reduced. Thus, we suggest that tyrosine or phenylalanine is also likely to interact with EA. The present study not only provides the clues for further studies concerning the mechanism of TRP channel regulation, but also building the pore region prediction model, thereby defining the protein structure of TRPC4 and TRPC5.
Recently the structure of mouse TRPC4 [20] and zebrafish TRPC4 [21] was released. There are two gates: the selectivity filter gate and the lower gate at the bundle crossing. The amino acid sequence of the pore region in TRPC5 is highly identical with TRPC4. Using the structure of TRPC4 to predict the location of three amino acids, K554, H594, and E598, we have located K554 between S5 and the pore helix, and H594 between the pore helix and S6 (Supplementary Fig. 4) Interestingly, E598 is located within S6. EA interacting with three amino acids might induce the conformational change in the pore helix and open the selectivity filter gate formed by N584 and G581. According to structural results of TRPC4, C549, and C554 form a disulfide bridge. We showed that C553 and C558 in TRPC5 as well as C549 and C554 in TRPC4 formed a disulfide bridge [2]. When the disulfide bridge was broken, the TRPC4/5 channels were translocated to the cytosol. Mutation of C553 (C549 in TRPC4) or C558 (C554 in TRPC4) to alanine or serine in TRPC5, the surface expression of channels was significantly decreased. Whole cell currents were not recorded at C553 or C558 mutants. In TRPC4, E555, located at the end of the pore turret formed a salt bridge with R556 of the adjacent domain. The mutation of K560 (corresponding to R556 of zebrafish TRPC4) to aspargine (N) decreased the EA-induced currents (Fig. 1), whereas the mutation of E559 (corresponding to E555 of TRPC4) to glutamine (Q) decreased the Cs+-induced currents with GTPγS in the internal solution [1].
The E598 residue in TRPC5 is a residue among residues responding to external La3+, Gd3+, Ca2+ and H+ [11422]. Semtner et al. [22] showed that H+ binds with E543 in TRPC5 activated by G proteins, whereas H+ binds with E595 in TRPC5 showing basal spontaneous activity. In the previous report, we suggested that E548, K554, and E598 as well as E543 and E595 are involved in the regulation of TRPC5 by H+ [1]. In addition, we showed that the effect of La3+ as well as H+ depended on extracellular cation condition, that is, mainly Na+ or Cs+ conditions [1]. Jung et al. [14] reported that E543, E595, and E598 are involved in the regulation of TRPC5 by La3+. The double mutation of Glu595/Glu598 in TRPC5 results in an acceleration of Mn2+ influx, an increase in relative Ca2+ permeability, a loss of potentiation by La3+, and a modification of channel inhibition. As Jung et al. [14] suggested, the possible explanation for the increase in the single-channel current of E595Q/E598Q could be due to attenuated block by a physiological cation, and the glutamates (E543, E595, and E598) form a negatively charged ring at the extracellular pore mouth, in tetramers, would act as “gatekeepers,” controlling the entry of cations into the pore. Duan et al . [20] showed that E559 also contributes to the negatively charged ring with help of C553–C558 disulfide bridge. The effect of E595Q/E598Q mutation on inhibition by La3+, the increase in single-channel current, and the increase in PCa/PNa also suggests that this site lies close to, or within, the permeation pathway [14]. Interestingly, E594 (corresponding to E598 of TRPC5) was found to be located within the S6 helix in TRPC4 [2021] as predicted [14]. In our hands, the response to EA or CCh in E595Q/E598Q mutant was similar to WT TRPC5, although the response of each mutant to EA decreased compared with WT. The degree of response to EA in mutants tends to follow the degree of TRPC5 activation by any agonist (Cs+, GTPγS, CCh). We investigated whether EA caused any permeability change to monovalent cations in WT TRPC5 or not. The EA-induced current showed the similar aspects for the current amplitude of Cs+ > Na+ > Li+ with that in CCh-induced currents (Supplementary Fig. 5). From the shift of the reversal potential at the various monovalent conditions, the sequence of Px was PCs > PNa > PLi (Supplementary Fig. 5). The permeability sequence was similar to TRPC4 and TRPC5 activated by GTPγS or CCh, whereas it was different from that in TRPC1/4 or TRPC1/5 heteromers activated by CCh. In this study, E595Q mutant responded to EA and showed EA-induced current showing typical double rectifying I/V curve, whereas E598Q did not. This result indicates that E598 is more important for EA binding than E595. R593N and E595Q responded to EA transiently (Fig. 2) while H594N did not respond (Fig. 1). Binding of EA to E598 might induce conformation change in the S6 helix and open the inner gate at the bundle crossing. K554, and H594 locates flanking pore helix, thus binding of both residues with EA might induce movement of pore helix and open the selectivity filter gate. R593, H594, and E595 seem especially important for EA action to be sustained.
EA activated TRPC1/TRPC5 heteromer with similar potency to TRPC4/5 homomer [923]. The sequence of EELQS in TRPC1 is different from HEFTE of TRPC4/5 whereas the sequence CVGIFCEQQ of TRPC1 is similar to CKGIRCEKQ of TRPC4/5 (Supplementary Fig. 4). These results suggest that EA mediates intersubunit interaction from two subunits of TRPC5. In addition, EA might activate TRPC1/5 heteromers by binding with both HEFTE of TRPC4/5 and CVGIFCEQQ of TRPC1 rather than HEFTE of TRPC4/5 and EELQS of TRPC1.
Rubaiy et al. [24] reported that A54, an EA analog, is a potent antagonist of EA, sharing the same interaction site, at homomeric TRPC5 channels and endogenous TRPC4/TRPC1-containing channels. A54 is also a strong enhancer of TRPC5 and TRPC4 channel activity when induced by Gd3+ and S1P (GPCR agonist), respectively. Rubaiy et al. [24] proposed that the EA and A54 data were explained by the two binding sites in close physical proximity: site 1 for binding without efficacy, and site 2 for binding with efficacy. In their model, EA would bind sites 1 and 2 and therefore, have a direct agonist effect. On the other hand, A54 would bind site 1 in the absence of a cofactor, thereby antagonizing EA and site 2 in the presence of the cofactor impairing the enhancing effect. The fact that cofactor Gd3+ binds to E595/E598 and A54 is a potent enhancer of the channel activity induced by Gd3+ suggests that E595/E598 is a part of the domain allowing A54 to bind with site 2 or site 2 itself. Additionally, it was reported in Chen et al. [25], that a residue, R593, placed in the pore loop close to the TRPC5 extracellular Gd3+ binding site, is critical to confer the sensitivity to GPCR-Gαq/11-PLC-dependent gating on TRPC5. GTPγS and GPCR agonists were shown to weakly activate the R593A mutant, while the addition of Gd3+ helped rescue the sensitivity of the mutant to GPCR-Gαq/11-PLC-dependent gating. These studies suggest that R593 and E595/E598 can have an important role as a domain that allows the binding of A54 with site 2, or site 2 itself. In our experiments, GTPγS and EA were shown to moderately activate TRPC5 R593N mutant (Fig. 1E) [1], and such a difference is hypothesized to be due to mutation of R593 to asparagine. Through a computer modelling based on rat TRPV1 structure, Chen et al. [25] showed the critical role of Y542, E543, R593, and E959 residues for an establishment of a functional Gd3+ binding site, and the participation of the Y541 residue in fine-tuning Gd3+ sensitivity of the TRPC5 channel. In our experiments, Y542F mutant didn't seem to express well, while Y541F showed moderate responses to EA (Fig. 1E) and GTPγS [1]. In the mTRPC5 structure, revealed by cryogenic electron microscopy (Cryo-EM) [26]. Cryo-EM structure of TRPC5 at 2.8 Å resolution reveals unique and conserved structural elements essential for channel function. As Duan et al. [26] and Li et al. [27] submitted, R593 interacts via a salt bridge with E598 and forms a hydrogen bond with the V590 side chain. These suggest the potential of R593 as the molecular fulcrum that can allow the GPCR-Gαq/11-PLC-powered gating force to efficiently transmit to mTRPC5's pore helix-loop [2627]. Such results of TRPC4 current activation by EA, but not by GPCR agonist in C549A/C554A double mutant [20] and in human TRPC5 C553S/C558S current (Supplementary Fig. 6), support our hypothesis that the transmission of the GPCR-Gαq/11-PLC-powered gating force to TRPC5's pore helix-loop may not be the only gating process. This EA's direct activation may possibly be another pathway to an opening of selectivity filter gate and the inner gate at the bundle crossing.
In conclusion, TRPC5 is activated by G protein and modulated by lipids, metal ions, and H+ [28]. EA interacts with at least three amino acids and induces the conformational changes in the selectivity filter gate and the inner gate at the bundle crossing, therefore opening and activating TRPC5 channel.



 XML Download
XML Download