

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
An imbalance between osteoblastic bone formation and osteoclastic bone resorption reduces bone mineral density and increases risk of fracture. In most cases, an increase in the ratio of osteoclast-to-osteoblast is due to estrogen deficiency during menopause [1]. In estrogen deficiency, receptor activator of nuclear factor-κB ligand (RANKL) is secreted from osteocytes and B cells [2]. Binding of RANKL to RANK on osteoclast progenitor cells leads to the recruitment of TNF receptor-associated factors (TRAFs) and subsequent activation of mitogen-activated protein kinases (MAPKs) and nuclear factor-κB (NF-κB) pathways [345]. These signaling pathways further induce nuclear factor of activated T cells c1 (NFATc1), a master regulator of osteoclast differentiation, and target the expression of genes such as matrix metallopeptidase 9 (Mmp9), osteoclast-associated receptor (Oscar), tartrate-resistant acid phosphatase 5 (Acp5), integrin beta 3 (Itgb3), and cathepsin K (Ctsk) [6]. Taken together, these findings suggest that the RANKL-RANK-TRAFs-NFATc1 axis is essential for the formation of mature osteoclasts after menopause.
Previously, we synthesized SPA0355 (1-methyl-3-[4-(2-phenoxazin-10-ylethoxy)phenyl]thiourea) and reported its anti-inflammatory effects in animal experimental models of rheumatoid arthritis [7], type 1 diabetes [8], hepatic ischemia-reperfusion injury [9], and allergic airway inflammation [10]. Biological actions of SPA0355 were related to the suppression of NF-κB and Janus kinase (JAK)/signal transducer and activator of transcription (STAT) pathways. Based on these observations, we speculated a possible beneficial effect of SPA0355 on ovariectomy (OVX)-induced bone loss in mice. Our results demonstrate that SPA0355 has preventive effects for osteoporosis by counteracting osteoclast-mediated bone loss and by stimulating osteoblast formation.
METHODS
Animals and experimental design
Pathogen-free 10-week-old C57BL/6 female mice were purchased from Orient Bio (Seoul, Korea). Mice were housed in a laminar flow cabinet with a 12 h light/dark cycle and maintained on standard laboratory chow ad libitum. Mice were anesthetized with ketamine (100 mg/kg) and xylazine (10 mg/kg) by intraperitoneal injection and subjected to either a sham operation or bilateral OVX. Mice were divided into four groups (n=5 per group): sham, vehicle (OVX+100 µl corn oil), and one of two doses of SPA0355 (OVX+10 or 50 mg/kg SPA0355 in 100 µl corn oil). The administration of vehicle or SPA0355 was started from day three after surgery and continued three times per week for five weeks. At the end of the study, femurs were fixed in 4% paraformaldehyde for µCT analyses. In toxicity study, SPA0355 was administered intraperitoneally at the dose of 50 mg/kg/day for seven consecutive days. This protocol was approved by the Institutional Animal Care and Use Committee at Chonbuk National University (permit number: CBNU 2015-87) and all work was performed in accordance with the National Institutes of Health guide for the care and use of Laboratory animals (NIH Publications No. 8023, revised 1978).
Dual energy x-ray absorptiometry (DEXA) and micro–computed tomography (µCT)
Five weeks after surgery, mice were anesthetized through ip injection of ketamine/xylazine, and their femur bone mineral densities (BMDs) were measured using a Lunar PIXImus densitometer (GE Lunar PIXImus Corporation Headquarters, Madison, WI, USA). Mice were then sacrificed, and structural changes in the femur were analyzed by SKYSCAN 1076 Micro-CT (Skyscan, Kontich, Belgium). The fixed femurs were scanned and reconstructed into three-dimensional images using NRECON software (version 1.6.1.5) and CT analyzer (version 1.10.0.1), both from SkyScan. Bone volume (BV), total volume (TV), percent bone volume (BV/TV), bone surface (BS), bone surface density (BS/TV), trabecular thickness (Tb.Th), and trabecular separation (Tb.Sp) were measured using the resident reconstruction program (Skyscan). A global algorithm was applied at a constant threshold for all specimens. The threshold was the intensity (gray value) that corresponded to ~50% of the average intensity of intact cortical bone. Trabecular lesions or metaphyseal cortical bone were analyzed in the distal metaphysis extending proximally 2.5 mm from the end of the growth plate.
Histological study
Fixed liver and kidney tissues were embedded in paraffin. Tissue sections (5 µm) were stained with hematoxylin and eosin (H&E) for light microscopy.
Hematological and biochemical assays
The plasma levels of hemoglobin (Hb), hematocrit (Hct), and the number of white blood cell (WBC) and platelet (PLT) were measured using automated hematology analyzer (XN-2000, Sysmex Co., Kobe, Japan). The serum levels of alanine aminotransferase (AST), aspartate aminotransferase (AST), blood urea nitrogen (BUN) and creatinine were measured using automated chemistry analyzer (ADVIA 2400, Siemens Healthcare Diagnostics Inc., Tarrytown, NY, USA).
Osteoclast differentiation
Bone marrow-derived macrophages (BMMs) were prepared as previously described [11]. BMMs were seeded at 2×106 cells/well in 6-well plates and cultured with macrophage colony-stimulating factor (M-CSF; Peprotech, London, United Kingdom; 30 ng/ml). After incubation for two days, cells were cultured in the presence of 10 ng/ml M-CSF and 30 ng/ml soluble RANKL (sRANKL, Peprotech). To detect formation and calculate the number of osteoclasts, BMMs were seeded at 5×104 cells/well in 48-well plates and cultured with M-CSF (10 ng/ml) and sRANKL (30 ng/ml) for four days. Osteoclasts were visualized by staining for tartrate-resistant acid phosphatase (TRAP) activity (Sigma-Aldrich, St. Louis, MO, USA). TRAP-positive cells were counted as osteoclasts.
Alizarin red S staining
MC3T3-E1 cells (5×104 cells/well) were cultured on 24-well plates and incubated in the differentiation medium (50 µg/ml ascorbic acid and 10 mM β-glycerophosphate) for 21 days. Then cells were fixed in 70% ice-cold ethanol and stained with 2% Alizarin red S. After washing with PBS, 10% cetylpyridinium chloride solutions was added to dissolve the calcium precipitates and absorbance was measured at 560 nm with a microplate reader.
Alkaline phosphate (ALP) staining and activity
MC3T3-E1 cells (5×104 cells/well) were cultured in 24-well plates and then incubated in the differentiation medium. On day 6, cells were fixed with 70% ice-cold methanol and stained with ALP staining solution (Sigma-Aldrich). For ALP enzymatic activity, cells were lysed at Day 6 with cell lysis buffer, and cell lysates were incubated with p-nitrophenyl phosphate (p-NPP) (Abcam, Cambridge, MA, USA). The color change of p-NPP to p-nitrophenol was measured at a wavelength of 405 nm.
RNA isolation and real-time quantitative RT-PCR (qPCR)
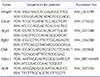
Total RNA was extracted from differentiated osteoclasts using an RNA Iso kit (TaKaRa, Tokyo, Japan). cDNA was prepared by reverse transcribing 1 µg of RNA with the first-strand cDNA synthesis kit (Applied Biosystems, Foster City, CA, USA). Primer sequences of genes used for quantification of mRNA by real-time PCR appear in Table 1. qPCR was performed using an ABI Prism 7900HT Sequence Detection System (Applied Biosystems).
Western blotting
Cell lysates (20 µg) were separated by 10% SDS-PAGE and transferred to PVDF membranes. After blocking with 5% nonfat dry milk, the membrane was probed with primary antibodies against p38, p-p38, p-AKT, AKT, p-ERK, ERK, JNK, p-JNK, p65, p-IκBα, IκBα (Cell Signaling, Beverly, MA, USA), or HSP90 (Enzo Life Sciences, Farmingdale, NY, USA). Membrane was incubated with goat anti-rabbit horseradish peroxidase–conjugated IgG (Santa Cruz Biotechnology, Santa Cruz, CA, USA) for 1 h. The proteins of interest on immunoblots were detected by using Las-4000 imager (GE Healthcare Life Science, Pittsburgh, PA, USA).
Luciferase
Transient transfections were performed using Lipofectamine 3000 (Invitrogen, Carlsbad, CA, USA) according to the manufacturer's instructions. Briefly, BMMs were transfected with plasmids encoding the NF-κB-luc reporter and the renilla luciferase reporter (pRL-TK-luc). The day following transduction, cells were treated with 30 ng/ml sRANKL, either with or without a 2 h pretreatment with 10 µM SPA0355. After 30 min, cells were harvested in reporter lysis buffer (Promega, Madison, WI, USA). Luciferase activity was determined in whole cell lysates using the Promega luciferase assay kit and is expressed relative to the renilla signal.
RESULTS
SPA0355 prevents OVX-induced bone loss
To determine whether SPA0355 could prevent bone loss induced by estrogen deficiency, mice were either sham-operated or ovariectomized with or without SPA0355 treatment. Five weeks after OVX, body weights were significantly increased compared to the sham operated mice (Fig. 1A). SPA0355 treatment at either dose (10 or 50 mg/kg) did not affect body weight significantly when compared to the untreated mice. In contrast to the body weight increase, the uterine weights decreased in the OVX mice when compared to the sham group (Fig. 1B). Again, SPA0355 treatment did not alter the uterine weight of OVX mice.
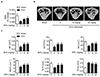
Next, we compared changes in bone mass. Ovariectomy caused a marked decrease of BMD in mice; however, treatment of SPA0355 (10 or 50 mg/kg) led to a significant increase of BMD compared to OVX mice (Fig. 2A). Consistently, µCT analyses also revealed bone loss in OVX mice: BV, BV/TV, BS, BS/TV, and Tb.Th were decreased, while Tb.Sp was increased in femurs compared to those of the sham group (Figs. 2B and C). However, the changes of these parameters were significantly attenuated in SPA0355-treated mice.
SPA0355 does not cause tissue damage
To investigate the potential side-effect of SPA0355, we administered SPA0355 daily for seven days. As shown in Table 2, SPA0355 at dose 50 mg/kg did not alter hematological parameters such as hemoglobin, hematocrit, white blood cell, and platelet. Plasma levels of AST, ALT, BUN, and creatinine (Table 2) and histological observations of liver and kidney (Fig. 3) showed no specific tissue damage by SPA0355 treatment when compared to the control mice.
SPA0355 inhibits osteoclast differentiation
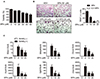
To determine whether SPA0355 inhibits osteoclast precursors and their differentiation into osteoclasts, the effect of SPA0355 (10 and 20 µM) was examined for RANKL-induced osteoclast formation in BMMs. SPA0355 at these concentrations was not toxic to BMMs during the four-day differentiation period (Fig. 4A). BMMs differentiated into osteoclasts in the presence of MCSF and sRANKL. Treatment of SPA0355 decreased the formation of TRAP-positive multinuclear cell formation (Fig. 4B). In the presence of 20 µM SPA0355, there was almost complete inhibition of osteoclastogenesis. We examined whether or not the inhibitory effect of SPA0355 on mature osteoclast formation is mediated via estrogen receptor. Treatment of BMMs with ICI 182,780, an estrogen receptor blocker, did not abrogate SPA0355 effect on osteoclast formation, indicating that SPA0355 action on the osteoclastogenesis is estrogen receptor-independent. We next examined the effect of SPA0355 on the expression of osteoclast marker genes. As shown in Fig. 4C, SPA0355 considerably decreased RANKL-induced mRNA levels of Mmp9, Oscar, Acp5, Itgb3, and Ctsk. SPA0355 also reduced RANKL-induced expression of Nfatc1, which has been reported to be a crucial regulator of osteoclastogenesis [6].
SPA0355 inhibits MAPKs, Akt, and NF-κB pathways in BMMs
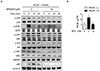
We examined the effect of SPA0355 on RANKL-induced signaling pathways in BMMs. RANKL rapidly induced the phosphorylation of p38, ERK, JNK, and Akt, and SPA0355 treatment inhibited all of these pathways (Fig. 5A). We also investigated whether SPA0355 could suppress the RANKL-induced NF-κB signaling pathway. SPA0355 inhibited RANKL-induced IκBα phosphorylation, NF-κB nuclear translocation, and NF-κB luciferase reporter activity (Figs. 5A and B). From these results, we could confirm that SPA0355 suppresses osteoclastogenesis by inhibiting RANKL-induced MAPKs, Akt, and NF-κB pathways.
SPA0355 stimulates osteoblastogenesis
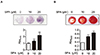
Because the balance between bone formation by osteoblasts and bone resorption by osteoclasts maintains adequate bone mass [1], we next questioned if SPA0355 has an osteogenic activity. We assessed the staining and activity of ALP, an early differentiation marker, in MC3T3-E1 cells. As shown in Fig. 6A, ALP staining and activity were significantly increased by SPA0355 at 10 and 20 µM concentrations. We also performed Alizarin red S staining to measure mineralization of the extracellular matrix as a marker for osteoblastic maturation. Treatment with SPA0355 significantly promoted bone nodule formation in a dose-dependent manner (Fig. 6B), indicating that SPA0355 acts on both initial and terminal steps of osteoblast differentiation.
DISCUSSION
The aims of this study were to elucidate the effects of SPA0355 on osteoclastogenesis and osteoblastogenesis in vitro and on OVX-induced bone loss in vivo. A previous study in our laboratory demonstrated that SPA0355 potently inhibits bone resorption in ankle joints of collagen-induced arthritic mice [7]. In order to examine the in vivo effects of SPA0355 treatment on bone turnover, we used an OVX-induced osteoporosis model. In agreement with earlier reports [1213], OVX resulted in a significant increase of body weight with concomitant decrease of uterine weight, indicating the success of the OVX. SPA0355 treatment did not have significant effects on body weight or uterine weight when compared to the control. Given that estrogen is the main hormone produced by the ovary and induces uterine growth [14] and that ICI 182,780 does not affect the action of SPA0355 on osteoclast formation, the osteo-protective effects of SPA0355 on an OVX-induced osteoporosis model do not depend on estrogen signaling.
Estrogen deficiency also accelerates osteoclast activity by indirect mechanisms. Estrogen withdrawal induces secretion of cytokines such as TNF-α, IL-6, IL-1, RANKL, and M-CSF from T cells, bone marrow stromal cells, osteoblasts, osteocytes, B cells. The cytokines promotes proliferation and differentiation of osteoclast precursors to mature osteoclast [215]. Especially, upon activation of RANK through binding of RANKL, the MAPKs, Akt, and NF-κB pathways are activated in osteoclasts [345]. A number of small molecules and natural products have proven to inhibit osteoclastogenesis and bone loss by inhibiting these pathways both upstream and downstream of RANKL signaling [1617]. Likewise, SPA0355 also inhibited MAPKs, Akt, and NF-κB pathways, which led to decreased expression of NFATc1 and its downstream genes, confirming the inhibitory effect of SPA0355 on RANKL-induced osteoclast differentiation.
In order to examine the in vivo effects of SPA0355 on bone remodeling, we used an OVX-induced osteoporosis model. The 10 and 50 mg/kg doses of SPA0355 used in this study are similar to those used in studies of rheumatoid arthritis [7] and streptozotocin-induced type 1 diabetes [8] and far greater than those used in studies of allergic airway inflammation [10] and hepatic ischemia-reperfusion injury [9]. It seems that doses of SPA0355 depend on the experimental period of the studies. OVX mice treated for five weeks with SPA0355 displayed improved trabecular femur BMD and other bone parameters (BV/TV, Tb.N, and Tb.Sp), which have been associated with bone loss after OVX. Unfortunately, no further improvement of any bone morphometric parameters measured by µCT scanning was observed with increase in SPA0355 dose from 10 to 50 mg/kg. The lack of dose-responsiveness may be due to a dose-saturation effect. Alternatively, the mild nature of the parameter changes may suggest that a substantially larger dose than the five-fold difference we used may provide a dose-effect. However, in this case, unexpected toxic side effects may develop.
The limitations of this study should be considered. First, we only evaluated osteo-protective effects of SPA0355 at the end of the treatment period. We also did not optimize the specific dosage range. Second, although menopause has considered as the single most critical risk factor for osteoporosis, aging also has adverse effects on skeletal homeostasis independent of estrogen deficiency [18]. Therefore, further study should investigate the effects of SPA0355 on age-related changes in the bone itself. Finally, the direct interaction target of SPA0355 to inhibit the MAPKs, Akt, and NF-κB pathways has yet to be clarified.
In summary, we demonstrated that SPA0355 suppressed RANKL-induced osteoclastogenesis in BMMs by inhibiting the activation of MAPKs (p-38, ERK, and JNK), Akt, and NF-κB and by suppressing the expression of NFATc1 (Fig. 7). This study discloses the beneficial effect of SPA0355 on OVX-induced bone resorption.




 XML Download
XML Download