

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Psoriasis and atopic dermatitis (AD) are chronic relapsing skin diseases characterized by dryness of the skin due to the impairment of the skin-barrier function1. Although the etiologies of both diseases have not been fully understood yet, studies on their pathogenesis have revealed that both are highly associated with the dysregulation of the immune system, mainly involving T cells. Psoriasis affects 2% to 3% of the whole population2. Its cause remains unknown even though genetic, environmental, and immunologic factors are known to be associated with the pathogenesis of psoriasis3. AD is characterized by severe pruritus and eczema. It has a prevalence of 10% to 20% in industrialized countries45. AD is highly associated with immune system imbalance, especially with T-helper (Th)2 immune responses6.
Vitamin C, also known as ascorbic acid, is a water-soluble ingredient found in fruits and vegetables. It has been traditionally utilized as an anti-oxidant and anti-aging agent7. However, vitamin C is unstable, easily oxidized, and decomposed into biologically inactive compounds8. In addition, intact vitamin C cannot easily penetrate the skin barrier due to its hydrophilic property. Therefore, a safer and stable delivery system for vitamin C needs to be developed by utilizing platforms such as microcapsules, nanospheres, or liposomes9. Choy et al.10 reported that unstable biological molecules can be successfully stabilized and effectively delivered to target tissues when they are encapsulated in a layered inorganic matrix of Vitabrid. Vitabrid is composed of the lamellar structure of hydrated zinc oxide (ZnO), which has been used in the management of many skin diseases because of its antioxidant, anti-inflammatory, and anti-bacterial effect11. Particularly, VitabridC12 is vitamin C incorporated into Vitabrid through the new technology of putting vitamin C in the layered complex of mineral carrier. It is composed of approximately 40-µm powdered particles. It formed into a nanoporous shell structure, which is coated with glyceryl monostearate. Glyceryl monostearate, emulsifier, can prevent the aggregation of vitamin C particles and increase skin's affinity values by hydrophobic modification of its surface.
The molecular mechanisms and therapeutic effect of VitabridC12, however, have not been clearly understood. In this study, we aimed to investigate the effect of VitabridC12 on skin diseases in psoriasis and AD mouse models by using imiquimod and Dermatophagoides farinae body (Dfb) extracts, respectively. Our results demonstrate that topical application of VitabridC12 significantly reduced skin lesions. The therapeutic action and effect of VitabridC12 on chronic inflammatory skin diseases was demonstrated in this study.
MATERIALS AND METHODS
All animal care was performed in accordance with the Guide for the Care and Use of Laboratory Animals (Washington DC, USA) and was approved by the Animal Care and Use Committee of The Catholic University of Korea (2014-0090-01).
Mice and materials
Female C57BL/6 and NC/Nga mice were purchased from Charles River Japan (Yokohama, Japan). These mice were all housed under conditions of controlled temperature (20℃~26℃), humidity (30%~70%), and lighting (lights on from 8 AM to 8 PM). Mice at 7 to 11 weeks of age were used for all the experiments and maintained in a specific pathogen-free barrier facility. VitabridC12 and Vitabrid were obtained from Hyundai IBT Ltd. (Seoul, Korea).
Establishment of imiquimod-induced psoriasiform dermatitis-like mouse models
Mice at 8 weeks of age daily received topical application of 62.5 mg of commercially available imiquimod cream (5%) (Aldara; 3M Pharmaceuticals, St. Paul, MN, USA) for 6 consecutive days. Control mice were daily treated similarly with a control vehicle cream (Vaseline Lanette cream; Fagron, Rotterdam, The Netherlands) for 6 consecutive days. VitabridC12 or Vitabrid were daily applied into the mouse back, followed 12 hours later by topical treatment with imiquimod. We performed experiments with five mice in each group every experiment. We repeated this experiment three times. Four experimental groups were as follows: control vehicle group (Vaseline was applied twice a day), imiquimod group (imiquimod and Vaseline were applied daily with 12-hour interval), Vitabrid group (imiquimod and Vitabrid were applied daily with 12-hour interval), and VitabridC12 group (imiquimod and VitabridC12 were applied daily with 12-hour interval).
Preparation of Dfb ointment and induction of AD-like mouse models
Dfb ointment, which is mixture of hydrophilic penetration and 5 mg of Dfb extract per gram hydrophilic petrolatum, was purchased from Biostir Inc. (Kobe, Japan). Hair on the upper back of the mice was removed with a clipper and a shaver 1 day before the experiment. Elicitation was performed by topical application of 100-mg Dfb ointment or ointment base (hydrophilic petrolatum) on the shaved dorsal skin and both surfaces of each ear. Barrier disruption was achieved by applying 150 µl of 4% sodium dodecyl sulfate on the shaved dorsal skin and both surfaces of each ear 3 hours before the Dfb ointment application. These procedures were repeated twice a week. We performed experiments with five mice in each group every experiment and repeated this experiment three times. Four experimental groups were as follows: control vehicle group (hydrophilic petrolatum was applied twice a week), Dfb group (Dfb ointment was applied twice a week), Vitabrid group (Dfb ointment was applied twice a week and Vitabrid was applied daily for 6 consecutive days. If Dfb ointment and Vitabrid needed to be applied on the same day, there was at least 12 hours' interval between Dfb ointment and Vitabrid treatments.), and Vitabrid C12 group (Dfb ointment was applied twice a week and Vitabrid C12 was applied daily for 6 consecutive days. Similar to Vitabrid, if Dfb ointment and VitabridC12 needed to be applied on the same day, there was at least 12 hours' interval between Dfb ointment and VitabridC12 treatments.).
Histological analysis
Skin lesions taken from the imiquimod- and Dfb-treated mice were fixed with 4% paraformaldehyde in phosphate-buffered saline (PBS) for 24 hours, washed with tap water, dehydrated with grade ethanol, and then embedded in paraffin. The paraffin blocks were cut in 4-µm-thick sections, mounted on glass slides, dewaxed, rehydrated with grade ethanol, and then stained with hematoxylin-eosin (H&E). Analysis was performed by using a microscope. All histological studies were performed in a blinded manner by two experienced dermatopathologists.
RNA isolation and quantitative real-time polymerase chain reaction
Total RNA was isolated from mouse skin by using the TRIzol reagent (Invitrogen, Carlsbad, CA, USA). The first cDNA strand was synthesized from 1-µg total RNA by using a QuantiTect Reverse Transcription Kit (Qiagen, Hilden, Germany). The QuantiTect primer assay (Qiagen) was used as primer sets for interleukin (IL)-1β, IL-4, IL-5, IL-6, IL-8, IL-13, IL-17A, IL-22, tumor necrosis factor (TNF)-α, CXCL1, CCL4, CCL17, CCL20, and thymic stromal lymphopoietin (TSLP). Glyceraldehyde-3-phosphate dehydrogenase messenger RNA was used as an endogenous control. Polymerase chain reaction (PCR) was performed by using Rotor-Gene 6000 (Corbett, Mortlake, Australia) and QuantiTect SYBR Green PCR Kit (Qiagen). The amplification program consisted of 1 cycle at 95℃ for 10 minutes, followed by 35 cycles at 95℃ for 20 seconds, 55℃ for 20 seconds, and 72℃ for 20 seconds.
Sodium dodecyl sulfate-polyacrylamide gel electrophoresis and western blotting analysis
Samples were separated by SDS-PAGE and transferred onto polyvinylidene difluoride membranes (Pall Corp., Port Washington, NY, USA). Membranes were blocked for 1 hour in a blocking buffer (5% skim milk in 25 mM Tris-HCl [pH 8.0], 150 mM NaCl, and 0.1% Tween 20) and incubated overnight at 4℃ with specific primary antibodies for target signaling. Blots were washed and incubated with horseradish peroxidase-conjugated secondary antibodies for 2 hours at room temperature, and the signals were detected by using radiography after reaction with an enhanced chemiluminescence detection kit (iNtRON Biotechnology, Seongnam, Korea).
Enzyme-linked immunosorbent assay
The amounts of total immunoglobulin E (IgE) in sera were determined by using sandwich enzyme-linked immunosorbent assay as previously described12. In brief, 96-well plates were coated with monoclonal anti-mouse IgE overnight at 4℃ and then further treated with 2% (w/v) bovine serum albumin dissolved in PBS containing 0.05% Tween 20 (PBS-T) for 2 hours at room temperature to block nonspecific binding. Next, serum samples were serially diluted in duplicated wells and incubated for 1 hour at room temperature. After 3 rinses with PBS-T, biotin-conjugated rat anti-mouse IgE Ab (LO-ME-2) was added to the wells, followed by the addition of streptavidin-peroxidase (Seikagaku Corp., Tokyo, Japan). After 3 rinses, the plates were developed by using a substrate solution containing 0.04% O-phenylenediamine dissolved in phosphate citrate buffer (pH 5.0). The reactions were terminated by the addition of sulfuric acid. The absorbance was read by using a microplate reader at 490 nm, and the amount of IgE isotype was calculated by comparison with the mouse IgE standard (Pharmingen, San Diego, CA, USA).
Statistical analysis
These results are representative of 3 independent experiments (n=5 for each group). Each data represents the mean value±standard error of three independent experiments. Statistical analyses were conducted with IBM SPSS Statistics ver. 19.0 for Windows (IBM Co., Armonk, NY, USA). Statistical analysis between multiple groups was performed using the one-way analysis of variance (ANOVA) (Kruskal-Wallis test) followed by Dunn's Multiple Comparison test. p-values <0.05 were considered statistically significant.
RESULTS
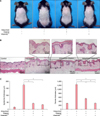
VitabridC12 alleviated imiquimod-induced psoriasiform inflammation in the mouse model
To examine the effects of VitabridC12 on psoriasis, psoriasiform dermatitis was induced on the back of the mice by the application of imiquimod daily for 6 days. Unlike the control group, the imiquimod group had induced skin swelling, which is the first hallmark of local skin inflammation (Fig. 1A). In addition, histopathological evaluation of the skin showed epidermal hyperplasia (acanthosis), hyperkeratosis, and increased dermal inflammatory cell infiltration in the imiquimod group compared to the control group (Fig. 1B). The thickness of the epidermis and dermis, and inflammatory cell infiltration significantly decreased in the Vitabrid or VitabridC12 group compared to the imiquimod group (Fig. 1C). Notably, the VitabridC12 group showed less inflammation than the Vitabrid group in the dermal thickness (p<0.05).
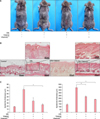
VitabridC12 alleviated the Dfb-induced AD-like inflammation in the mouse model
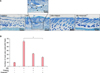
To examine the effects of Vitabrid C12 on the AD murine model, Dfb ointment was applied on the back of Nc/Nga mice twice for 3 weeks. Changes in the skin thickness after 3 weeks are shown in Fig. 2. Apparent acanthosis and edematous skin swelling occurred after the application of Dfb ointment in NC/Nga mice compared to the control group. Histological examination revealed an increased inflammatory response, including dense dermal infiltration of leukocytes, epidermal hyperplasia, and hyperkeratosis in the Dfb ointment-treated mice skin. As we expected, VitabridC12 was effective in decreasing these inflammatory responses (Fig. 2B). In the VitabridC12 group, the thicknesses of the epidermis markedly decreased, compared to the Dfb group (p<0.05, Fig. 2C). Interestingly, in toluidine blue staining, the mast cells more significantly decreased in the VitabridC12 group than in the Dfb control group (p<0.05, Fig. 3).
VitabridC12 suppressed the expressions of the inflammatory mediators in psoriasiform dermatitis
VitabridC12 was found to be effective in inhibiting the development of imiquimod-induced cutaneous inflammation in mice skin. Next, we performed quantitative real-time PCR analysis in mice skin to examine the mRNA expression levels of proinflammatory mediators. Similar to the histological changes, real-time PCR analysis revealed that the mRNA expression levels of proinflammatory cytokines such as IL-1β, IL-6, IL-8, TNF-α, IL-17A, and IL-22 increased in the imiquimod-treated mouse ears, whereas these mRNA expression levels decreased significantly in the VitabridC12 group (Fig. 4). VitabridC12 inhibited the mRNA expressions of IL-17A and IL-22, the signature cytokines of the Th17 cell lineage. Furthermore, treatment with VitabridC12 significantly hindered the mRNA expression levels of the other inflammatory chemokines such as CXCL1, CCL17, and CCL20, which are highly elevated in psoriasis131415.
VitabridC12 suppressed the expressions of the inflammatory mediators and IgE in the AD mouse model
We also investigated whether VitabridC12 downregulated the production of cytokines and chemokines in the Dfb-induced skin lesions of the NC/Nga mice. Quantitative real-time PCR analysis revealed that the mRNA expression levels of T cell-specific cytokines such as IL-4, IL-5, and IL-13 increased in the Dfb-treated mice ears, whereas VitabridC12 inhibited these mRNA expression levels. Compared to Dfb group, VitabridC12 is significantly inhibiting the mRNA expressions of TSLP, IL-4, IL-13 and CCL4 (p<0.05, Fig. 5A). Furthermore, VitabridC12 significantly downregulated the mRNA expression of TSLP, IL-4, and CCL4 more effectively than Vitabrid (p<0.05, Fig. 5A).
Then, we tried to investigate whether VitabridC12 could modulate the serum IgE levels in the Dfb-induced mice. The total IgE levels in the sera obtained from the mice are shown in Fig. 5B. The IgE levels increased in the Dfb-treated mice. The VitabridC12-treated mice displayed relatively lower levels of total serum IgE than Vitabrid-treated mice (p<0.05).
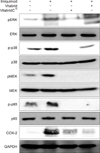
VitabridC12 downregulated the MAPK and NF-κB signaling pathways
In addition, we examined whether VitabridC12 could inhibit mitogen-activated protein kinase (MAPK) signaling pathways that play key roles in inflammatory signaling in mammalian cells1617. Nuclear factor (NF)-κB p65 is the active subunit of NF-κB. Overactivation of NF-κB signaling leads to inflammatory disorders18. COX-2 is an inducible enzyme that converts arachidonic acid into prostaglandins and is involved in various inflammatory diseases19. VitabridC12 inhibited phosphorylation of MAPK/ERK kinase (MEK), extracellular signal-regulated kinase (ERK), and p38 in the imiquimod-treated mice. Moreover, VitabridC12 hindered phosphorylation of NF-κB p65 and reduced the expression levels of COX-2 in the imiquimod-treated mice (Fig. 6). Thus, we might speculate that VitabridC12 could downregulate cutaneous inflammation via inhibition of the MAPK kinase and NF-κB pathways.
DISCUSSION
VitabridC12 is a novel compound composed of a glyceryl monostearate -coated, layered vitamin C-Vitabrid complex. Vitamin C has long been regarded as a potent antioxidant agent20. However, because of its instability and the difficulty in delivering it into the dermis, many studies have been conducted to find stable compounds of and new delivery systems for vitamin C. Vitabrid is a hydrated ZnO in the lamellar structure, which causes minimal skin irritation10. ZnO also helps the skin restore disturbed skin-barrier function and enhance wound healing with its antimicrobial effects2122. It has also an anti-inflammatory effect by reducing neutrophil chemotaxis in psoriatic patients23. Because VitabridC12 is coated with hydrophobic glyceryl monostearate, the innovative substance penetrates into the skin more efficaciously than other vitamin compounds. Whereas vitamin C can be easily destroyed within 3 to 4 hours of exposure to air, VitabridC12 consistently releases vitamin C for longer than 12 hours. Furthermore, as VitabridC12 exchanges ions with perspiration (NaCl) from sweat glands and CO2 from the air, it can help in the absorption of vitamin C in the deep skin layer.
However, the biological and pharmacological activities of VitabridC12 are not well understood, specifically how it works in skin inflammatory diseases. Therefore, we investigated whether VitabridC12 could modulate skin inflammation in psoriasis and AD mouse models by using imiquimod and Dfb ointment, respectively. Topical application of VitabridC12 suppressed imiquimod-induced skin thickness and dermal inflammatory cell infiltration. The results suggested that treatment with VitabridC12 could exert significant anti-inflammatory effects on cutaneous inflammation. In addition, Vitabrid C12 significantly suppressed the expressions of proinflammatory mediators, including TNF-α, IL-1β, IL-6, IL-8, IL-17A, IL-22, CXCL1, CCL17, CCL20, and COX-2, which were elevated in psoriatic skin lesions2425262728293031.
Previous studies showed that in psoriatic skin lesions, the expression levels of ERK, JNK, and p38 were increased in the MAPK pathway163233. Therefore, we investigated the MAPK and NF-κB signaling pathways3435363738. Our study demonstrated that VitabridC12 could reduce the activities of p38, ERK, and MEK. While multiple signaling pathways, including NF-κB, are dysregulated in psoriatic epidermis39, VitabridC12 treatment reduced the phosphorylation of p65, a component of the NF-κB pathway. Our results showed that the therapeutic effect of VitabridC12 on the expression and production of proinflammatory mediators is associated with the inhibition of the MAPK kinase and NF-κB signaling pathways.
To study the effect of VitabridC12 on AD, we induced AD in NC/Nga mice by applying Dfb ointment. Our results showed that VitabridC12 efficiently suppressed the infiltration of inflammatory cells, including mast cells, and decreased the levels of the Th2 subset cytokines. Elevated serum IgE level is observed in most AD patients40. VitabridC12 also reduced the serum IgE level in the NC/Nga mice. AD keratinocytes secrete TSLP, which activates dendritic cells to prime naive T cells to produce IL-4 and IL-1341. In chronic AD, IL-5 is involved in eosinophil development and survival42. CCL4, also known as macrophage inflammatory protein 1β, is a CC chemokine with specificity for CCR5 receptors43. Serum CCL4 level is significantly elevated in patients with AD. Topical application of VitabridC12 significantly reduced AD symptoms and the levels of AD-related markers, including IL-4, IL-5, IL-13, CCL4, and TSLP. Sivaranjani et al.44 suggested that antioxidants such as Vitamin C may be helpful in the treatment of AD. We suggested that stable Vitamin C with Vitabrid may have synergic effect to AD.
The results of this study show that VitabridC12 reduce the cutaneous inflammation in psoriasis and AD model mice via modulating the complex network of cytokine-mediated signaling cascades. Moreover, VitabridC12 effectively inhibited diverse signaling pathways, including MAPK and NF-κB signaling. These results also propose that VitabridC12, a stable form of vitamin C with ZnO in the lamellar structure, has an enhanced ability to control skin inflammation. In conclusion, we propose the use of VitabridC12 as an effective treatment option for psoriasis and AD.
In summary, we observed that topical application of VitabridC12 decreased the number of inflammatory cells and the expression levels of diverse markers, thus reducing psoriasis or AD-like symptoms in the mice in this study. The effect of VitabridC12 might be associated with NF-κB and MAPK signaling pathways. Collectively, our results suggest that Vitabrid C12 is a promising candidate as a treatment option for management of psoriasis and AD.


 XML Download
XML Download