

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Asthma is a chronic bronchial inflammatory disease associated with several processes, including hyper-reactivity of the bronchi and local inflammation/thickening of the bronchial walls leading to mucus formation and bronchoconstriction.123 According to the World Health Organization (WHO) estimates, nearly 235 million individuals suffer from asthma worldwide and its incidence, constantly increasing, is often underrated due to misdiagnosis. Though some cases of asthma, especially developing in childhood period, can be remitted and disappear, the disease is not usually curable for life because about 10% of asthma patients do not respond to currently available therapies4 Asthma control is based on a daily background treatment (anti-inflammatory drugs, inhaled corticosteroids, or anti-leukotrienes) and an emergency treatment (fast-acting bronchodilators).567 Difficulty in developing efficient therapeutic strategies may reflect the fact that the pathogenesis of asthma is poorly understood.
There are several cytokines and chemokines that are involved in the pathophysiology of asthma. Cytokines bind to membrane-specific receptors of their target cells and trigger signal transduction pathways that alter gene expression in these target cells.8910 Currently, cytokines are considered therapeutic targets, and even some specific anticytokines, such as interleukin (IL)-5 antibodies (mepolizumab or reslizumab), have already been approved as a treatment option for severe asthma.1112
The role of IL-13, particularly in severe asthma, has been confirmed. In bronchial biopsy specimens of severe asthmatic patients, there is a large number of inflammatory cells with increased expression of mRNAs encoding IL-13.1314. In vivo animal experiments have shown the critical role of IL-13 in the development of asthma.1516 In vitro, IL-13 showed a significant effect on various cellular processes within the airway and correlated with the clinical aspects of airway diseases.17 Specifically, IL-13 is found to induce apoptosis and fibrosis in lung fibroblasts18 and cooperates with alveolar macrophages and dendritic cells to promote inflammation and chemokine release.19 In addition, IL-13 positively regulates collagen type-1 in airway fibroblasts in asthma.20 Therefore, establishing innovative therapeutic tactics to target this critical cytokine will be vital for therapeutic purposes.2122
The detection of key microRNAs (miRNAs), a class of small, noncoding, single-stranded RNAs that regulate gene expression, may be useful for setting up therapeutic strategies for asthma, via probable therapeutic modulation using miRNA mimics or inhibitors. In previous studies, miR-155 was found to be downregulated in many pathological conditions, including cancer and cardiovascular disease, and decreased miR-155 expression was involved in the pathogenesis of asthma.2324 Mechanistic studies about the role of miR-155 in asthma pathophysiology have previously been reported using in vitro and in vivo models. For example, miR-155 was found to be essential for T(H)2-mediated allergen-induced eosinophilic inflammation in the lung. In addition, miR-155 is a critical regulator of type 2 innate lymphoid cells and IL-33 signaling in experimental models of allergic airway inflammation. However, the potential role of miR-155 in asthma pathophysiology and its underlying mechanism remain to be determined. In addition, to the best of our knowledge, studies investigating the regulatory effect of IL-13 on miRNAs in asthma pathogenesis are scarce and ill-defined, especially regarding miR-155 and its functional role.
Therefore, the aim of the present study was to determine the regulatory effect of IL-13 on the expression of miR-155 and to explore the role of miR-155 in the proliferation and migration of IL-13-induced bronchial smooth muscle cells (BSMCs) and the underlying molecular mechanism involved.
MATERIALS AND METHODS
Cell culture
Human BSMCs were obtained from Lonza (Walkersville, MD, USA). The M199 culture medium added with 10% new born calf serum (ThermoFisher Scientific, Carlsbad, CA, USA), 0.5 µg/L epidermal growth factor, and 2 µg/L fibroblast growth factor was employed for cell cultivation in a humidified 5% CO2 atmosphere at 37℃. Once cell growth reached 70%–80% confluence, it was stopped for 48 hours by transfer in serum-free media containing a 50:50 mix of F12/DMEM enhanced with 10 mL/L ITS Premix (BD Bioscience, Bedford, MA, USA). Cells were then treated with different concentrations of IL-13 for 24 hours or left untreated. IL-13 was purchased from Sigma-Aldrich (St. Louis, MO, USA).
Transfection
The MISSION® hsa-miR-155 mimic (HMI0254) or hsa-miR-155 hairpin inhibitor (HSTUD0254) were obtained from Sigma-Aldrich corporation. The human transforming growth factor (TGF)-β-activated kinase 1/MAP3K7-binding protein 2 (TAB2) small interfering RNA (siRNA) (h) was purchased from Santa Cruz Biotechnology. BSMCs were seeded onto 96-well plates, and once cells reached 50%–80% confluence, they were transfected with hsa-miR-155 mimic (10 nM)/hsa-miR-155 inhibitor or TAB2 siRNA following the manufacturer's instructions. Cells were harvested after 72 hours for subsequent analyses.
In silico analysis of predicted miRNA targets
The online bioinformatics' tool TargetScan (Whitehead Institute for Biomedical Research, Cambridge, MA, USA; http://www.targetscan.org) was exploited to uncover candidate binding sites of miR-155 and the corresponding seed sequences of target transcripts.
Luciferase reporter assay
The miR-155 binding sites from the 3′-untranslated region (UTR) of TAB2 or mutant 3′-UTR of TAB2 were cloned into the control pGL3 downstream of the open reading frame (ORF) encoding the Firefly luciferase at the XbaI restriction site (GeneChem, Montreal, QC, Canada). Recombinant clones with the 3′-UTR regions inserted in the sense orientation were selected. BSMCs were cultured in 96-well plates. Aliquots of 10 nM of miR-155 mimic or control miRNA were cotransfected with 100 ng of the pGL3-TAB2 3′-UTR wildtype or mutant TAB2 plasmid DNA into BSMCs using Lipofectamine 2000. After transfection for 48 hours, luciferase assays were performed using the Dual Glo Luciferase Assay System™ (Promega, Madison, WI, USA) kit. The cells of each well were lysed using the lysis buffer (PLB 5X) for 15 minutes with stirring. The cell extracts were then transferred to a 96-well microplate and the bioluminescence intensities were determined using the Xenius XL™ luminometer (SAFAS corporation, Clifton, NJ, USA). The relative luciferase activity was standardized against Renilla data (ratio luciferase activity/renilla activity). The values were then quantitated from the control sample considered to be 100%. The experiments were conducted in triplicate.
Quantitative polymerase chain reaction (PCR)
Total RNA containing miRNA was extracted and enriched for the using miRVana isolation protocols (Ambion; ThermoFisher Scientific). The miScript II RT Kit was used for reverse transcription of total RNA, including miRNA into cDNA. The extracted RNA was quantified with a NanoDrop device (ThermoFisher Scientific) and kept in a refrigerator at −70℃ pending subsequent analysis. The expression level of miR-155-5p was assessed using the Taqman MicroRNA Reverse Transcription Kit (Applied Biosystems, Foster City, CA, USA). The quantitative real-time polymerase chain reaction (qRT-PCR) experiments were performed using the hsa-miR-155 Primers (QIAGEN, Germantown, MD, USA), TAB2 Sense (5′-CGATCAGCTGTTGCGAGCGCTGCAC-3′) and anti-sense (5′-GTGCAGCGCTCGCAACAGCTGATCG-3′) primers on the CFX96 Touch™ Real-Time PCR system (Bio-Rad, Hercules, CA, USA). The small nuclear RNA U6 was used as endogenous control for miR-155 while glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was the housekeeping gene used for TAB2. The ΔΔCt approach was applied to estimate the relative gene expression levels. The experiments were performed in triplicates. The following settings were used for PCR amplification experiments: 30 seconds at 95℃, followed by 45 cycles at 95℃ for 5 seconds and 58℃ for 34 seconds.
Cell proliferation assays
Three-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay was used to measure the effect of miR-155 on the proliferation of human BSMCs. Briefly, cells transfected with miR-155-5p mimic/inhibitor, negative controls, or untransfected cells were stimulated with IL-13 (80 ng/mL) and cultured for 72 hours. Next, 20 µL of 5 mg/mL MTT reagent was added to the culture, followed by additional incubation for 4 hours. After replacement of culture supernatant by 150 µL of dimethyl sulfoxide (DMSO) for solubilization of the formazan crystal, the absorbance at 570 nm was measured by using spectrophotometry.
Cell migration assays
The migration of BSMCs was evaluated by the Boyden chamber migration assay using 8-micron Transwell filters. BSMCs transfected with miR-155 mimic/inhibitor, negative controls, or untransfected cells were placed in the upper chamber at a density of 4,000 cells/well. Migration of BSMCs was stimulated by addition of different concentrations of IL-13 to the lower compartment. Next, cells were left in the chamber for 6 hours in terms of which cells that had migrated through the membrane were dyed, photographed, and counted with the help of the Axiovert 135 microscope. Assays were performed in triplicate.
Western blot analysis
Cells were treated with RIPA buffer at 4℃ for 15 minutes for cell lysis. Then, BCA Protein Assay kit was used to determine protein concentration of cell lysates. After protein purification on 10% sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE), aliquots were transferred on the polyvinylidene difluoride (PVDF) membrane and blocked with 5% bovine serum albumin (BSA) in tris-buffered saline containing 0.1% Tween 20 (TBST) at ambient temperature for 1 hour. The membranes were incubated at 4℃ overnight with anti-TAB2 antibody (ab172412; Abcam, Cambridge, MA, USA) or anti-GAPDH antibody (ab9483, Abcam) as a loading control. After incubation, the blots were washed 3 times with TBST and incubated in presence of horseradish peroxidase (HRP)-conjugated secondary antibodies prior to visualization with the Pierce ECL Western Blotting Substrate (ThermoFisher Scientific). Densitometry analysis was performed using the Image J software (National Institutes of Health, Bethesda, MD, USA).
Statistical analysis
All statistical analyses were conducted using Graphpad Prism software version 6.01 for windows (GraphPad Software, La Jolla, CA, USA). The data are expressed as mean±standard deviation. Differences between the groups were assessed by one-way analysis of variance (ANOVA) followed by the Bonferroni multiple comparison tests or two-way ANOVA, with Bonferroni multiple comparison posttests. A P value of <0.05 was considered statistically significant.
RESULTS
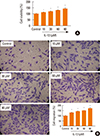
Treatment with IL-13 induces proliferation and migration of BSMCs
In order to reliably explore the effects of IL-13 related pathways on the proliferation and migration of BSMCs, IL-13-treated cells were subjected to MTT cell proliferation and Transwell migration assays. The results indicated that IL-13 treatment significantly induced the proliferation of BSMCs in a dose-dependent manner (Fig. 1A). Similarly, in Transwell assay, the migration of BSMCs was significantly increased following IL-13 treatment, dose-dependently (Fig. 1B).
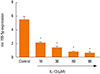
Expression of miR-155-5p is down-regulated in IL-13-induced BSMCs
To identify the role of miR-155-5p in the regulation of BSMC phenotype, miR-155-5p expression was examined in BSMCs treated with different concentrations of IL-13 which is known as a key cytokine driving asthma pathogenesis. The qRT-PCR experiments indicated that miR-155 expression was significantly and dose-dependently down-regulated in IL-13-induced BSMCs compared with the control untreated cells (Fig. 2).
Overexpression of miR-155-5p inhibits proliferation and migration of IL-13-induced BSMCs
The transfection of BSMCs with miR-155-5p mimic or inhibitor significantly promoted or inhibited the expression of miR-155. Similarly, overexpression of miR-155-5p inhibited the proliferation of BSMCs, while the inverse effect was found for miR-155-5p inhibitor (Fig. 3A). Meanwhile, during the migration assay, we observed that inhibition of miR-155-5p was accompanied by increased number of migrated cells, whereas transfection with miR-155-5p mimic showed a remarkable hindrance of BSMC migration ability (Fig. 3B).
TAB2 is a direct target of miR-155
To elucidate the mechanisms responsible for the regulation of BSMC phenotype by miR-155, we used bioinformatics to identify the target transcripts of miR-155. TAB2 was predicted as a potential miR-155 target (Fig. 4A). Therefore, we assessed TAB2 expression at protein and mRNA levels in BSMCs induced by different concentrations of IL-13. The mRNA expression of TAB2 was significantly and dose-dependently increased in IL-13-induced BSMCs compared with untreated cells (Fig. 4B). Western blot analysis showed that TAB2 protein level was equally significantly and dose-dependently increased in IL-13-induced BSMCs compared with untreated cells (Fig. 4C). Furthermore, using the luciferase reporter assay, we found that miR-155-5p significantly inhibited luciferase activity in BSMCs transfected with the vector harboring the wild-type 3′-UTR TAB2, while no significant difference was recorded with cells transfected with mutant 3′-UTR vector (Fig. 4D). These observations indicated that TAB2 was a direct target of miR-155-5p. Inhibition of miR-155-5p with the inhibitor resulted in the up-regulation of TAB2 protein expression in IL-13-induced BSMCs (Fig. 4E). On the contrary, miR-155 mimic led to decreased TAB2 protein levels. Neither miR-155 inhibitor nor mimic affected TAB2 mRNA expression (Fig. 4E). These results indicated that miR-155-5p is a posttranscriptional regulator of TAB2 and that miR-155-5p negatively regulated TAB2. These results suggested that overexpression of miR-155-5p inhibits the proliferation and migration of IL-13-induced BSMCs by suppressing TAB2.
Silencing of TAB2 mimics the effects of miR-155-5p on BSMCs
To evaluate possible involvement of miR-155-5p/TAB2 in IL-13-induced proliferation and migration of BSMCs, we determined the effects of TAB2 siRNA on the effects of BSMCs transfected with miR-155 mimic. MTT assay showed that TAB2 siRNA mimicked the inhibitory effects of miR-155 mimic on IL-13-induced proliferation of BSMCs (Fig. 5A). Similar effects were recorded on cell migration in Transwell migration assay (Fig. 5B). The present data indicated that miR-155-5p-mediated inhibition of TAB2 is involved in the inhibitory effect of mir-155-5p on the proliferation and migration of BSMCs. The results equally indicated that the IL-13/miR-155-5p/TAB2 pathways could be involved in the proliferation and migration of BSMCs, which can probably be implicated in the pathogenesis of asthma and other pulmonary diseases.
DISCUSSION
Cytokines are known as the main drivers of inflammatory diseases, such as asthma. TH2 cytokines (IL-4, IL-5, and IL-13) are related to disease activity, symptom score, eosinophilia of the airways, and bronchial hyper-reactivity. They increase after an allergenic stimulation and decrease with the corticoid treatment. Similar to IL-4, IL-13 stimulates the production of immunoglobulin E (IgE) by B lymphocytes, activates monocytes and macrophages, and increases mucus secretion by goblet cell hyperplasia.2526 Recent publications have highlighted the involvement of IL-13 in asthma,2728 but the molecular mechanism involved in this process is poorly studied. In asthma, IL-13 is reported to induce mucus production.293031 Additional studies also indicate important functions of IL-13 in airway smooth muscle cells, such as gene regulation, cell proliferation, and migration.323334 Specifically, IL-13 was found to promote contractility of BSMCs by up-regulating RhoA protein.35 In addition, IL-13 was found to induce BSMC proliferation by upregulating CysLT1 receptor IL-13 in response to LTD4.36 In the present study, we observed that IL-13 dose-dependently induced the proliferation and migration of BSMCs, thus confirming its role in bronchial asthma and the necessity to further explore its underlying mechanism.
Studies on the cross-talk between IL-13 and miRs in the pathophysiology of asthma, especially the phenotypic changes of BSMCs, are scarce and the underlying mechanism is ill-defined. In mouse in vivo studies, a set of miRs that are involved in the response to allergen attacks have been discovered and some of them, involved in the regulation of IL-13 and TH2 which are key components in asthmatic reactions, have been proved promising for pre-miR and anti-miR manipulations.3738 Other studies have identified miRs that are implicated in bronchial smooth muscle hyperresponsiveness and proliferation.404142 Up to now, the most frequently studied miR in the regulation of asthma molecular and cellular processes is miR-155, which is found to be up-regulated in asthma.3943 Previous studies have demonstrated that miR-155 participates in allergic airway inflammation through regulating the transcription factor PU.1.44 In vivo experiments indicated that miR-155 deficiency contributes to allergic airway inflammation improvement by modulating Th2 responses and ATP-/P2R-induced activation of dendritic cells in mice.45 An additional study documented that miR-155 promotes the proliferation of Th cells in allergic asthma by down-regulating cytotoxic T-lymphocyte-associated protein 4 (CTLA-4).46 All these findings suggested miR-155 to be a potential therapeutic target for allergic asthma. However, the molecular and cellular mechanism via which miR-155 exerts its action in asthma is not fully elucidated and needs further clarification. The present study was to scrutinize the mechanisms involved in the cross-talk between miR-155-5p and IL-13 and their action on the proliferation and migration of BSMCs, which could have beneficial implications in the understanding and treatment of pulmonary diseases, especially asthma. The main results were as follows: 1) IL-13 induced proliferation and migration of BSMCs dose-dependently; 2) IL-13 dose-dependently decreased the expression of miR-155-5p in IL-13-induced BSMCs; 3) Overexpression of miR-155-5p inhibited the proliferation and migration of IL-13-induced BSMCs; 4) TAB2 was a direct target of miR-155-5p and silencing of TAB2 mimicked the effects of miR-155-5p. Previous studies have shown that in human macrophages, miR-155 directly targets IL13Ralpha1 and reduces the levels of IL13Ralpha1 protein, leading to diminished activation of signal transducer and activator of transcription 6 (STAT6) and demonstrated that miR-155 affects the IL-13-dependent regulation of several genes (SOCS1, DC-SIGN, CCL18, CD23, and SERPINE) involved in the establishment of an M2/pro-Th(2) phenotype in macrophages. Our study, to the best of our knowledge, is the first to report the involvement of the IL-13/mir-155/TAB2 axis in the proliferation and migration of BSMCs. These findings corroborated, along with a previous study, that miR-155 is a negative regulator of TAB2 in mesenchymal stem cells (MSCs).47 A direct functional role of TAB2 in asthma has not been reported previously, but previous studies demonstrated its involvement in the activation of inflammatory pathways.474849 Our findings also provided evidence that IL-13-inhibits TAB2 expression via up-regulating miR-155-5p in BSMCs, which suggest its role in asthma inflammation.
Our work presents some limitations. IL-13-induced asthma hyperresponsiveness may be one of the key features in asthmatics, in which airway smooth muscle contraction plays a critical role. Nevertheless, in the present study, we did not check increased Ca++ oscillations or cell contraction in IL-13-treated BSMCs; thus, it is hard to precisely define the function of miR-155-overexpressed BSMCs. Further studies are needed to clarify this aspect.
We conclude that overexpression of miR-155-5p may inhibit the proliferation and migration of IL-13-induced human BSMCs by suppressing TAB2. In addition, the results suggest that modulation of the IL-13/mir-155/TAB2 pathway could be a potential strategy in the treatment of allergic asthma.



 XML Download
XML Download