

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Penicillin treatment failure was first reported soon after the introduction of penicillin administration in the 1940s; dissemination of penicillin resistance has made this antibiotic useless since the 1950s [1]. The mechanism of penicillin resistance prevalent during the 1960s and mid-1970s is associated with mutations in chromosomal genes (mainly penA gene) encoding for penicillin binding proteins (chromosomally-mediated penicillin resistant N. gonorrhoeae, CMRNG) [12]. Although CMRNG resulted in clinical treatment failure, administration of high-dose penicillin remained effective against CMRNG until a new resistance mechanism emerged in 1976 [3]. This novel mechanism resulted from the acquisition of a plasmid-located β-lactamase gene (blaTEM-1) exhibiting high-level resistance to penicillin (penicillinase-producing N. gonorrhoeae, PPNG). Wide dissemination of PPNG ultimately provoked the usage of extended-spectrum cephalosporin antibiotics, including ceftriaxone and cefixime, which were effective against the TEM-1 β-lactamase with a narrow hydrolytic spectrum [45]. However, there have been frequent recent reports of new β-lactamase variants sharing common single nucleotide polymorphisms (compared with TEM-1).
TEM-135, which has a single amino acid change (M182T) compared with the TEM-1 sequence, was first reported in 2005 [6], and its frequency and genetic diversity have been consistently evaluated in many countries. The clinical significance of TEM-135 remains unclear, as there are no major differences in the minimal inhibitory concentrations (MICs) of penicillin and extended-spectrum cephalosporins; however, the emergence of TEM-135 could signify the introduction of other TEM β-lactamases with an extended β-lactam hydrolytic spectrum through mutation or plasmid acquisition under the high selective pressure of cephalosporin usage. We aimed to determine the prevalence of TEM-135 and compare the characteristics of TEM-1- and TEM-135-producing N. gonorrhoeae isolated in Korea from 2005 to 2015.
METHODS
1. N. gonorrhoeae isolates
A total of 804 isolates were collected over 11 years (2005–2015) from 40 diverse hospitals and primary-care clinics in Korea to test for the presence of the TEM-135 gene. This study was approved by the Institutional Review Board of the Human Research Protection Center in the Yonsei University Health System (4-2016-0359). Transgrow medium was used as the transport medium to minimize the loss of viable N. gonorrhea isolates, and modified Thayer-Martin medium (Becton-Dickinson, Cockeysville, MD, USA) was used for colony growth and incubation. Bacterial identification was confirmed by Gram-stain and the Vitek system (bioMerieux, Marcy l'Etoile, France) or by matrix-assisted laser desorption/ionization-time of flight (MALDI-TOF) mass spectrometry (Bruker Daltonics, Bremen, Germany). All isolates were collected for the national surveillance program for gonococci supported by the Korean Centers for Disease Control and Prevention. They were stored at −70℃ until the number of isolates collected was sufficient for analysis. Molecular analysis was performed.
2. Antimicrobial susceptibility testing and examination of β-lactamase production
The antimicrobial susceptibility of all isolates to the antibiotics (penicillin G, tetracycline, ceftriaxone, spectinomycin, ciprofloxacin, nalidixic acid, and azithromycin) was determined using the CLSI disk diffusion method [7]. MIC was calculated using the agar dilution test according to the CLSI guidelines [8] for all antibiotics except azithromycin, for which the European Committee on Antimicrobial Susceptibility Testing (EUCAST) cut-off values were used [9]. The median and 90% MIC values (MIC50 and MIC90) and range were evaluated for the three groups of isolates: non-PPNG, PPNG with TEM-1, and PPNG with TEM-135.
3. DNA extraction and mismatch amplification mutation assay polymerase chain reaction
DNA was extracted from PPNG bacterial suspensions, using the QIAquick gel extraction kit (Qiagen, Hilden, Germany). Mismatch amplification mutation assay (MAMA) PCR was performed to detect the blaTEM-135 allele, and TEM PCR was performed to identify both blaTEM-1 and blaTEM-135 [10]. The complete blaTEM sequences of PPNG isolates were confirmed by Sanger sequencing based on a comparison between the TEM-135 reference sequence (GenBank accession number: GQ896333) and the PCR products.
4. Molecular epidemiology of TEM-135 isolates using N. gonorrhoeae multiantigen sequence typing (NG-MAST) and plasmid typing
To evaluate the molecular epidemiology of the TEM-135 isolates, NG-MAST was performed by sequencing the porB and tbpB genes; allele numbers and sequence types (STs) were assigned using the NG-MAST website (http://www.ng-mast.net), as previously described [11]. For plasmid typing, multiplex PCR was performed on all isolates, as previously described, using primers BL1, BL2, BL3, and BL4 [12].
RESULTS
1. Detection and trends of PPNG isolates with TEM-135
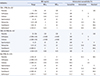
PPNG isolates accounted for 11.6% (N=93) of the 804 isolates, with patterns fluctuating by collection year (Fig. 1). Only 72 PPNG isolates (77.4%) were available for further molecular investigation because of loss or deterioration of the isolates during the storage period. These 72 isolates were subjected to full sequence analysis to determine the presence of TEM-1 or TEM-135; 22.2% (N=16) were shown to harbor TEM-135. The first TEM-135 isolate was identified in 2006, and the number of TEM-135 isolates has continuously increased since 2012. Although no variant types other than TEM-1 and TEM-135 were found, all isolates possessed a silent mutation, c.18C>T, which does not change the original amino acid (Proline) compared with the previously reported blaTEM-135 reference sequence (GQ896333; Table 1).
2. Antimicrobial susceptibility of PPNG isolates with or without TEM-135
Of the 72 PPNG isolates, 52 (38 TEM-1 and 14 TEM-135 isolates) were randomly selected (mainly because of strain availability) for antimicrobial susceptibility testing with seven antibiotics. The proportions of susceptible isolates for the seven antibiotics were similar between the TEM-1 group and TEM-135 group, whereas the non-PPNG group showed a different pattern (Table 2). Comparison of the MIC50 and MIC90 (determined using the agar dilution test) of the three groups (non-PPNG, PPNG with TEM-1, and PPNG with TEM-135) showed that the MIC50 and MIC90 of ceftriaxone and cefixime were significantly lower in the TEM-135 than in the other groups (all values ≤0.008 µg/mL). Additionally, the MIC50 and MIC90 of azithromycin in the TEM-135 group were also lower than those in the TEM-1 group (MIC50=0.06 µg/mL and MIC90=0.25 µg/mL for the TEM-135 group). No significant changes in antimicrobial resistance trends were observed in the TEM-1, TEM-135, and non-PPNG groups (Supplemental Data Table S1).
3. Determination of molecular epidemiological characteristics by NG-MAST and plasmid typing
The STs of 72 PPNG isolates were determined by NG-MAST in order to elucidate the molecular epidemiology of plasmids possessing certain types of penicillinases; 59 different STs were identified, indicating heterogeneous epidemiological distribution (Fig. 2). The STs bearing TEM-135 were dispersed throughout the genetic tree (generated based on por gene sequences using the unweighted pair group method with arithmetic mean [UPGMA] approach), except for two small clusters (cluster A and B; Fig. 2). Plasmid typing of all PPNG isolates demonstrated that the African plasmid type was the most common type, present in 51/72 isolates (70.8%), followed by the Toronto/Rio (N=13, 18.1%) and the Asian (N=8, 11.1%) plasmid types. However, among the TEM-135 isolates, the Toronto/Rio plasmid type was predominant (N=13, 81.3%), followed by the Asian (N=2, 12.5%) and the African (N=1, 6.2%) plasmid types. ST3965 isolates all possessed TEM-1 on an African type plasmid, except for one isolate with TEM-135 on a Toronto/Rio type plasmid. In cluster A, five TEM-135 isolates (designated with black dots and bold red numbers; blue box, Fig. 2) appeared to be closely related based on close genetic distances (determined by NG-MAST), the predominance of the Toronto/Rio type (established by plasmid typing), and recent isolation (within the last three years). Four isolates in cluster B (Fig. 2) harbored three Toronto/Rio plasmids and one Asian plasmid. Interestingly, the Asian and African plasmids bearing TEM-135 have appeared recently (in 2012, 2014, and 2015).
DISCUSSION
The emergence of PPNG was one reason for the use of extended-spectrum cephalosporins, including ceftriaxone and cefixime; the appearance of TEM-135 could constitute another turning point in gonococcal treatment through the evolution or introduction of novel β-lactamases to N. gonorrhoeae. Following the first report of TEM-135 in Japan, the reported prevalence of TEM-135 ranged from 9.4% to 57.8% according to country [101314151617]. The average prevalence of TEM-135 in our study was 22.2% over a period of 11 years (2005–2015). Although the prevalence trends showed fluctuations, a pattern of continuous increase in the proportion of TEM-135 is observed in the last four years (2012–2015).
The antimicrobial susceptibility patterns of ceftriaxone, cefixime, azithromycin, and tetracycline in PPNG with TEM-135 differed from those in PPNG with TEM-1. Although all PPNG isolates were susceptible to ceftriaxone and cefixime, the MIC50 and MIC90 of both antibiotics were much lower in PPNG with TEM-135 (both values ≤0.008 µg/mL) than in PPNG with TEM-1 (0.03 µg/mL and 0.12 µg/mL, respectively). The rate of azithromycin-susceptible isolates was 100% in TEM-135 strains, but 60% in TEM-1 strains. The MIC50 of tetracycline in TEM-135 and TEM-1 PPNG was 0.5 µg/mL and 4 µg/mL, respectively, and the rates of resistance were 30.8% and 78.1%, respectively. Although a previous study conducted in England utilizing a similar approach discovered statistically higher ciprofloxacin and tetracycline MIC values in TEM-135 strains [14], our results do not support such trends. However, they may suggest clonal relatedness consisting of isolates highly susceptible to ceftriaxone and cefixime that can be discriminated from TEM-1 PPNG, despite the high selective pressure of ceftriaxone usage in Korea.
This hypothesis can be partially supported by our phylogenetic and plasmid analyses. The phylogenetic tree based on partial porB sequences revealed that some of the TEM-135 isolates appear to be closely related in terms of genetic distances and recent isolation within the last three years (Cluster A and B; Fig. 2). However, the other genetically dispersed TEM-135 PPNG isolates were also highly susceptible to ceftriaxone and cefixime, which may suggest that TEM-135 PPNG isolates originated in a low ceftriaxone selective pressure environment (similar to tetracycline-resistant N. gonorrhoeae [TRNG] that has increased recently in Korea [18]), compared with TEM-1 PPNG, which is less susceptible to ceftriaxone and cefixime.
The predominance of the Toronto/Rio plasmid type in TEM-135 PPNG revealed in our study is in line with previous epidemiological studies conducted in other countries [10151619], although only one study from England reported the predominance of the Asian plasmid type [14]. Therefore, we assume that a specific plasmid (Toronto/Rio) has been distributed worldwide. The ceftriaxone and cefixime MIC values of isolates with an Asian plasmid type (0.06 µg/mL, respectively) were higher than the values of isolates with other plasmid types; however, the number of isolates was too small to draw any definitive conclusions.
A recent report identified four novel TEM alleles from globally collected strains [19]; we did not identify any new TEM alleles other than TEM-1 and TEM-135. Interestingly, all isolates in this study had a thymidine instead of cytosine at nucleic acid position 18 compared with the previously reported blaTEM-135 reference sequence from Thailand (GenBank accession number: GQ896333). However, a comparison of our sequences with N. gonorrhoeae TEM-1 and TEM-220 sequences revealed that thymidine at position 18 is common (Table 1). Moreover, the blaTEM-135 sequence in Salmonella enterica subsp. enterica serovar Typhimurium, in which TEM-135 was first identified, was identical to that of all our TEM-135-harboring isolates. This sequence is also identical to the sequences of the first TEM-135 isolates reported in Japan [17]. Thus, the TEM-135 PPNG isolates from our study might be more closely related to the original Japanese TEM-135 isolates than to the previously reported reference strain. Furthermore, the clonal relatedness of TEM-135 strains only became prominent in the last three years, suggesting the possibility of foreign acquisition.
In conclusion, the proportion of TEM-135 PPNG has continuously increased during the past four years and most of the isolates are highly susceptible to ceftriaxone and cefixime despite the high selective pressure of ceftriaxone usage in Korea. The recent increase in TEM-135 PPNG proportion is associated with clonal spread; however, we hypothesize that most of the isolates have originated from relatively low ceftriaxone selective pressure environments. Moreover, the difference at position 18 of the TEM-135 sequence can be interpreted as the presence of at least two clonal complexes, one of which is thought to be associated with the Japanese sequence, even though direct connection cannot be proved. The possibility of foreign import requires rigorous follow-up to control the foreign acquisition of novel resistance and continuous monitoring of TEM-135 to elucidate whether TEM-135 constitutes a step towards evolutionary change of resistance to prevent the dissemination.



 XML Download
XML Download