

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
The granule cells (GCs) of the cerebellum are the most numerous neurons of the entire central nervous system and lie within the internal granular layer (IGL), which is located below a monolayer of Purkinje cells (PCs). Above the PC layer lies the molecular layer, where the GCs form parallel fibers that make excitatory synapses on PC dendrites. GC precursors disperse from the upper rhombic lip (URL) in a rostral direction over the dorsal surface of the cerebellum to form the external granular layer (EGL). Altman and Bayer have extensively studied the development of the EGL in rats and reconstructed the EGL dispersal pathway over the cerebellum (Altman, 1972; Altman & Bayer, 1985; 1997). They found that the EGL proliferates from the rhombic lip at E17 in an anterodorsal direction. Medial (or vermal) EGL cells disperse rostrally and lateral (or hemispheric) cells disperse medially.
A paired homeobox DNA binding protein, Pax6, is expressed in GCs and their precursors. A mutation in the Pax6 gene was found in aniridia patients (Hanson et al., 1993). These patients do not develop irises. Mice and rats with a null mutation in Pax6 were identified and dubbed Small Eye. Animals heterozygous for the mutation have smaller eyes than their normal littermates. Homozygous animals do not develop eyes or nasal cavities, and they die shortly after birth (Hill et al., 1991). It was found that Pax6 was crucial for eye development (Hanson & Van Heyningen, 1995). Pax6 has also been found to play a major role in the development of pancreatic islets (Beimesche et al., 1999). In the central nervous system, Pax6 has been implicated in axon pathfinding, neuron migration and the formation of axon tracts and projections (Mastick et al., 1997; Kawano et al., 1999). In the cerebellum, Pax6 is expressed in both the EGL and the IGL (Engelkamp et al., 1999; Yamasaki et al., 2001). Cerebella of homozygous Small Eye mice have a narrower vermis and lack foliation when compared with wild-type littermates. Though dispersal of the EGL appeared normal in Small Eye, GCs failed to migrate transversely and completely failed to migrate inwards (Engelkamp et al., 1999). Yamasaki et al. found that this is due to a cell autonomous defect in polarization in mutant GCs. Because of this defect, mutant GCs do not develop a leading process and migrate from the EGL in random directions (Yamasaki et al., 2001).
Research has been done on the migratory pathways of GCs for over a century. However, previous methods, involving either cresyl violet (Nissl) staining or 3H-thymidine birthdating, were not conclusive (e.g. Altman & Bayer, 1997; Beierbach, 2001). When specific antibodies were used, either only postnatal tissue was labeled (Karam et al., 2001), or, when embryonic tissue was labeled, the studies focused on Pax6 expression in the whole brain was studied or focused on aspects of cerebellar development separate from EGL dispersal (Engelkamp et al., 1999; Jensen et al., 2004). Therefore, the aim of this study was to investigate the dispersal pattern of the EGL in more detail. By labeling during early embryonic development with an antibody against Pax6, we found that Pax6-immunoreactive neurons in the developing cerebellum are indeed early inward-migrating granule cells.
Materials and Methods
Mice
Pregnant CD1 mice were purchased from Charles River Laboratories (Montréal, Canada). The breeding date was defined as E0. Mice were housed according to local governmental and institutional animal care guidelines and fed ad libitum. Dams were sacrificed by cervical dislocation at pregnancy dates E11 to E18, and the embryos were removed. The embryonic brains were dissected, immersion fixed in 4% paraformaldehyde (PFA) (EMD Biosciences, San Diego, USA), 0.9% NaCl and 0.1 M phosphate buffer (pH 7.4) (PBS) (Sigma, St. Louis, USA), and stored at 4℃ until further processing.
Antibodies
The following primary antibodies were used:
- rabbit anti-Pax6 (used diluted 1:2000, Chemicon Inc. Temecula USA), which recognizes granule cells (Engelkamp et al., 1999; Yamasaki et al., 2001)
- mouse anti-NeuN (1 : 2,000, Chemicon Inc.), which recognizes differentiated granule cells (Weyer & Schilling, 2003)
- rabbit anti-Pax2 (1 : 1,000, Chemicon Inc.), which recognizes stellate and basket cell precursors (Maricich & Herrup, 1999)
Sectioning and immunohistochemistry
Embryonic cerebellums were obtained from pregnant mice. Embryonic cerebellums were quickly washed with 0.9% NaCl in 0.1 M phosphate buffer (PBS: pH 7.4) followed by 4% paraformaldehyde in 0.1 M phosphate buffer (pH 7.4) for 48 hours. The cerebella were then cryoprotected through a series of buffered sucrose solutions: 10% (2 hrs), 20% (2 hrs) and 30% (overnight). Series of 40 µm thick transverse sections were cut through the extent of the cerebellum on a cryostat and collected for free-floating immunohistochemistry. Briefly, tissue sections were washed thoroughly, blocked with 10% normal goat serum (Jackson ImmunoResearch Laboratories, West Grove, PA) and then incubated in 0.1 M phosphate buffered saline (PBS) containing 0.1% Triton-X and the primary antibody for 16~18 hours at 4℃. Secondary incubation in horseradish peroxidase (HRP) conjugated goat anti-rabbit or HRP conjugated goat anti-mouse antibodies (all diluted 1 : 200 in PBS; Jackson ImmunoResearch Laboratories, West Grove, PA) was subsequently performed for 2 hours at room temperature. Diaminobenzidine (DAB, 0.5 mg/ml) was used to visualize the reaction product. Finally, sections were dehydrated through an alcohol series, cleared in xylene and coverslipped with Entellan mounting medium (BDH Chemicals, Toronto, ON, Canada).
Cerebellar sections were also processed for fluorescent immunohistochemistry. Tissue sections were washed, blocked in PBS containing 10% normal goat serum (Jackson ImmunoResearch Laboratories, West Grove, PA), then incubated in both primary antibodies overnight at room temperature, rinsed, and incubated for 2 hours at room temperature in a mixture of Alexa 546-conjugated goat anti-rabbit Ig and Alexa 488-conjugated goat anti-mouse Ig (Molecular Probes Inc., Eugene, OR), all diluted 1 : 2,000. After several rinses in 0.1 M PBS, sections were coverslipped in non-fluorescing mounting medium (Fluorsave Reagent, Calbiochem, La Jolla, CA).
BrdU injection and immunohistochemistry
Dams were injected with 100 µl 18.2 mg/mL bromodeoxyuridine (BrdU, Sigma) on pregnancy day 15 and sacrificed after a survival period of 10 minutes according to the protocol of Miller and Nowakowski (1988). To label granule cells cells, dams pregnant for 10 days were injected with 100 µl 18.2 mg/ml BrdU and sacrificed on pregnancy day 15. The embryonic cerebella were removed, fixed, sectioned and stored as described above. Prior to immunohistochemistry, sections were treated with HCl (1 N, Titristar EMD Sciences) for 30 minutes at RT. Mouse anti-BrdU (1 : 5,000, G3G4, DSHB, Iowa, USA) was used to detect incorporated BrdU.
Photomicrographs were captured with a SPOT cooled color digital camera (Diagnostic Instruments Inc.) mounted on a Zeiss microscope and assembled in Adobe Photoshop (version 9). The images were cropped and corrected for brightness and contrast but not otherwise manipulated.
Results
Inward migration from the dorsal surface of the cerebellum before external granular layer (EGL) dispersal
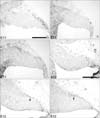
Cerebella at embryonic age E11 to E18 were sectioned horizontally or sagittally and stained for Pax6. No cells immunoreactive for Pax6 are observed in the cerebellum at E11 (Fig. 1A~C). At E12, a rostrodorsal group of cells immunoreactive for Pax6 is seen in both lateral and paramedial sections (Fig. 1E, F). At E12, a single layer of Pax6-immunoreactive cells on the dorsal surface of the cerebellum is observed that connects the rostral group of Pax6-immunoreactive cells to the upper rhombic lip (URL) (Fig. 1D~F).
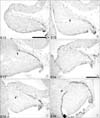
The dorsal layer of Pax6-immunoreactive cells increases to 3 cells thick by E13 (Fig. 2B) and to 6 cells thick by E14 (Fig. 2C), and is now referred to as the EGL. In medial sections, no dorsal layer of Pax6-immunoreactive cells is observed until E14 (Fig. 2D), suggesting that paramedial and lateral EGL dispersal may start earlier than medial EGL dispersal. In lateral and paramedial sections, Pax6-positive cells within the cerebellum are located more rostrally than the leading edge of the EGL (Fig. 4E, F).
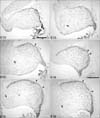
At E15, the rostral groups of Pax6-positive cells are most distinguishable (Fig. 3A~C). At E16, the inward migrating rostral group of Pax6-immunoreactive cells is less well defined in paramedial sections (Fig. 3E). At E17, we observed an area in the center of the cerebellum in paramedial sections that contains no Pax6-positive cells (Fig. 4B). Interestingly, this is not the case in the lateral sections, where a group of Pax6-immunoreactive cells can still be distinguished at the rostral end of the cerebellum at E17 and E18 (Fig. 4C, F). This suggests that Pax6 expression is down-regulated in at least the medial nucleus.
Identification of the early inward migrating Pax6-immunoreactive cells
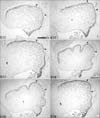
The rostral Pax6-positive group of cells within the cerebellum is most distinguishable at E15. To determine the identity of these cells, dams were injected with a non-lethal dose of BrdU on E10, sacrificed on E15, and sections were immunofluorescently double labeled for Pax6 and BrdU. In paramedial sections, many, but not all, of the rostral Pax6-positive group of cells are also immunoreactive for BrdU (Fig. 5B, C). It general, it appeared that most double labeled cells are more ventrally located than cells immunoreactive for Pax6 alone. Two possible explanations are that the latter cells were born either before BrdU injection, or after, when no more BrdU was available. These data suggest that these cells were not actively dividing at the time of BrdU injection.
To determine the identity of the other Pax6-immunoreactive cells within the cerebellum that appeared to have migrated inwards from the EGL, E15 sagittal and horizontal sections were immunostained with several cell type-specific markers. When sections are stained for Pax2, immunoreactive somata are seen scattered from the rostral end of the VZ to the dorsocaudal part of the cerebellum (Fig. 5A), which corresponds with the known migration pattern of stellate- and basket cell precursors (Maricich & Herrup 1999).
When E15 sagittal sections are immunolabeled for NeuN, an antigenic marker of mature GCs (Weyer & Schilling, 2003), no immunoreactive profiles are seen in the cerebellum, suggesting that the Pax6-positive cells within the cerebellum are not comprised of mature GCs (data not shown). To investigate when the Pax6-positive cells within the cerebellum mature into GCs, E16, E17 and E18 sagittal sections were double immunolabeled for Pax6 and NeuN. NeuN expression is first observed at E16 in a group of cells extending from the center of the cerebellum to the dorsal surface of the posterior lobe. All cells that are immunoreactive for NeuN are also immunoreactive for Pax6 (Fig. 5D). At E17 and E18, the number of double-positive cells increases (Fig. 5E, F) but remain in the same location, i.e. in a cluster extending from the center of the cerebellum into future lobules VII and VIII.
Discussion
In this study we have shown that Pax6 is expressed in both EGL cells and their derivatives, as well as in migrating deep cerebellar nuclei (DCN) cells. When Pax6 expression was studied more closely, details about the migrating DCN, the EGL and its dispersal were observed.
It is widely accepted that the inward migration of granule cells from the EGL does not begin until birth. Previous studies have shown that granule cells migrate primarily through raphes between postnatal days (P)0 and P7. After P7, no raphes are observed and granule cells migrate inwards en masse (Luckner et al., 2001; Karam et al., 2001). However, in this study we found that postmitotic Pax6-positive cells migrate from the EGL as early as E14, and double immunostaining with various cell markers suggested that these cells are likely GCs. These early inward migrating GCs did not appear to migrate in raphes but rather were scattered throughout the whole cerebellum. After E14, granule cells continued to migrate from the EGL. At E15, the GCs were found from the EGL to the ventral neuroepithelium, indicating that they had migrated past the location of the future IGL. From E16 to E18, Purkinje cells became more concentrated below the EGL, and the number of GCs seen in this layer decreased. At E18, GCs began to concentrate below the future PC layer in the future IGL, and no GCs were seen in the PC layer in most parts of the cerebellum.
Studies have sought to determine what signals GCs to migrate inward at specific locations. Cadherins and ephrins, two families of cell-adhesion molecules, are thought to play roles in determining the location of raphes. Raphes were found at the borders of PC clusters expressing different types of cadherins, Eph receptors and ephrin ligands (Karam et al., 2001; Luckner et al., 2001). In our study, GCs appeared to freely migrate inward until the PCs formed a layer. Therefore, we hypothesize that the inward migration of GCs is mechanically blocked by PCs between E16 and E18. In future studies, we plan to label P5 to P7 sections for Pax6 or RORα to investigate whether the start of the massive inward migration of GCs coincides with the transition from a multiple cell layer to a single layer of PCs.
Our finding that the early inward migrating cells from the EGL mature within the cerebellum led us to hypothesize that these cells may not be post-mitotic when they leave the EGL. Our BrdU data show that very few BrdU immunoreactive cells were observed At E15 (E15 injection and sacrificed 10 minutes later), and no Pax6 immunoreactive cells within the cerebellum were immunoreactive for BrdU. This suggests that Pax6-immunoreactive cells within the cerebellum were not proliferating at the time of BrdU exposure. Therefore we conclude that early inward migrating GCs, like the postnatally inward migrating GCs, are postmitotic, even though they do not express NeuN until E16. Taken together, our findings provide novel views on specific stages of granule cell dispersal and migration.
 XML Download
XML Download