

 PDF
PDF ePub
ePub Citation
Citation Print
Print


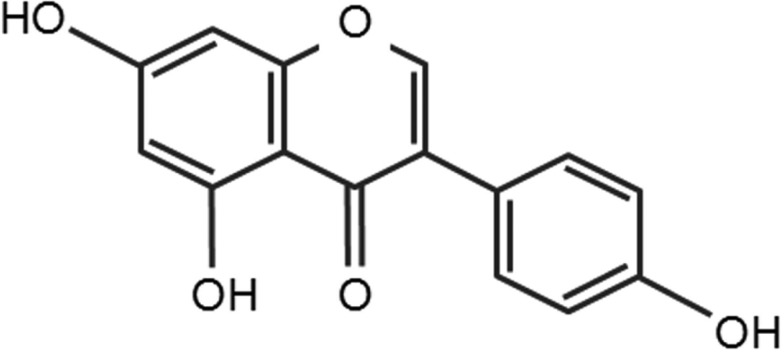
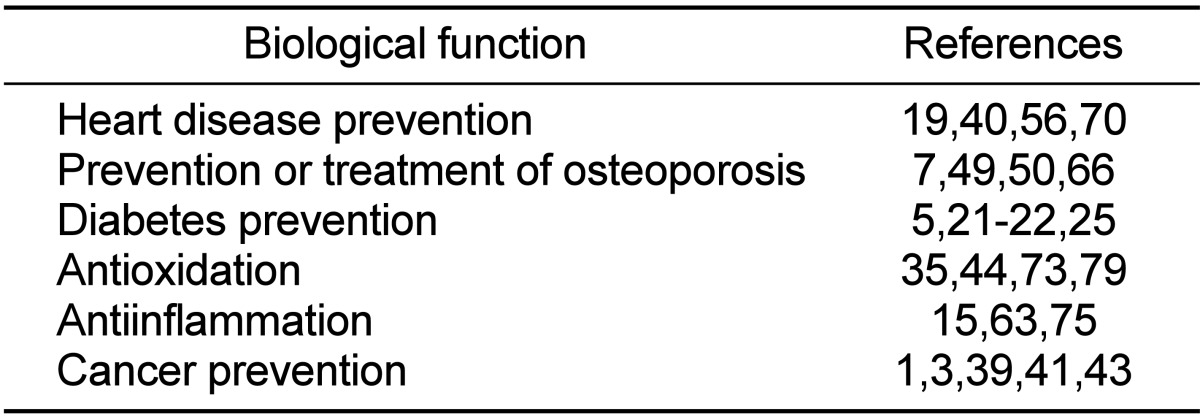
Genistein (5,7-Dihydroxy-3-(4-hydroxyphenyl) chromen-4-one) belongs to the category of isoflavones which are ketone or non-ketone polyhydroxy polyphenol compounds (Figure 1). This is mostly derived from a leguminous plant [13] and plant secondary metabolites fulfilling many functions such as plant pigmentation, UV filtration, and symbiotic nitrogen fixation. In addition, it is well known as one of phytoestrogens that are biologically active phenolic compounds derived from plants and have structures similar to the principal mammalian estrogen, 17β-estradiol (E2) [65]. Due to their structural similarity to E2, they have an estrogenic as well as an antiestrogenic effect and therefore affect many biological functions associated with endogenous estrogen actions. Genistein was first isolated in 1899 from the dyer's broom, Genista tinctoria, which is a species of flowering plant of the family Fabaceae. Therefore, the name of genistein was derived from a generic name of the plant from that it was first separated. Nowadays, soy is known to be the most abundant source of genistein. Many scientific studies have found that genistein may be helpful for preventing heart disease [19,40,56,70] and preventing or treating osteoporosis [7,49,50,66]. Additionally, multiple in vitro and animal researches have confirmed the antidiabetic properties of genistein while there are still limited data on the effects of genistein consumption in humans with diabetes [5,21,22,25]. Genistein also has other useful biological effects such as antioxidation [35,44,73,79] and antiinflammation [15,63,75]. There are also many evidences suggesting potential benefits of genistein in cancer prevention and treatment [1,3,39,41,43]. These diverse biological functions of genistein were summarized in Table 1.
Chemopreventive properties against cancers of genistein has emerged from epidemiological observations indicating that the incidence of some cancers including breast and prostate cancers is much lower in Asia, where diets are rich in soyfoods, than Western countries [1,2,30,59]. This epidemiological evidence may be further supported by the fact that breast cancer rates among Asian-Americans who migrated to the US are considerably higher than the rates in Asia [20,81]. The reason of increased breast cancer incidence in Asian-Americans was partially explained by the reduced intake of soyfoods after migration to the US [81]. These findings have shown a significant association between soyfood intake and risk of breast cancer and further evidence of chemoprevention effect of genistein, a major isoflavone in soybeans. Since 1990 when the chemopreventive efficacy of genistein became a possibility on the basis of epidemiological studies, enormous amount of scientific research has been conducted using diverse experimental tools [48]. In vitro cellular models adopting various cancer cell lines, in vivo animal models adopting the implantation of cancer cells or the administration of carcinogens, and further clinical studies have been used to examine and establish the possibility of genistein as a chemopreventive agent.
This article will review the chemopreventive activities against diverse cancers of genistein elucidated in animal studies which are usually conducted in preclinical phase to verify the underlying mechanisms elucidated in in vitro cellular models and to provide a foresight for clinical studies. Especially, as animal studies have advantages in designing the experiments to suit the goals, they imply diverse information on the anticancer activity of genistein. Although the case control or clinical studies targeted at humans have more important issues, clear consensus regarding the anticancer effect of genistein has not yet emerged due to the limitations including the difficulties in determining sample size, test period, and dietary or exposure measurement et al [48]. In this review, in vivo anticancer activities of genistein identified in animal studies will be discussed with being divided into its effects on carcinogenesis or cancer initiation and cancer development or growth.
Anticancer Activities of Genistein on Carcinogenesis
Carcinogenesis is a creation of cancer by the result of a process in which normal cells are transformed into cancer cells. The process ultimately reprograms a cell to undergo uncontrolled cell division, thus forming a malignant mass [27]. Carcinogenesis is caused by the mutation of genetic material of normal cells [12]. The genetic mutations leading to carcinogenesis are triggered by diverse factors including carcinogens, radiation, and viral transformation [9]. Among these factors, carcinogens are particular substances directly involved in causing cancer, which include natural carcinogens such as aflatoxin B1 produced by fungus Aspergillus flavus [34,67] and industrial carcinogens such as dioxine and dioxine-like compounds, benzene, benzo[a]pyrene, and vinyl chloride [46].
In animal experiments studying cancer, 7,12-Dimethylbenz[a]anthracene (DMBA), a polycyclic aromatic hydrocarbon (PAH), has been extensively used as a model carcinogen [51]. As an immunosuppressor and powerful organ-specific laboratory carcinogen, DMBA serves as a tumor initiator mostly in skin and mammary gland. Therefore, the inhibitory effects of genistein on carcinogenesis have been actively examined in DMBA induced cancer models using rodents. In the DMBA rat mammary cancer model, injections of genistein to female Sprague-Dawley (SD) rats on days 2, 4, 6, 16, 18, and 20 postpartum suppressed DMBA-induced mammary tumorigenesis. Because DMBA was administered on day 50 postpartum in these experiments, neonatal and prepubertal exposures of genistein are considered to be protective against mammary carcinogenesis [37,38,53]. These protective effects of genistein at the prepubertal period have been linked to decreased numbers of terminal end buds (TEBs) and increased numbers of lobular structures, leading to the effect of providing less sites for malignant transformation [28]. Additionally, it has been reported that the elimination of targets for mammary malignant transformation by prepubertal genistein exposure was attributed to genistein-induced upregulation of BRCA1, a tumor suppressor gene participating in DNA damage repair processes and cell differentiation, in the mammary gland [8]. The diet supplemented with genistein (111 ppm) also inhibited DMBA-induced carcinogenesis in female CD rats by reducing DMBA-DNA binding and DNA-adduct formation in mammary tissues [72]. Another study using DMBA and testosterone propionate (TP) - induced rat carcinogenesis models also provided evidence that genistein may have potential as a chemopreventive agent against carcinogenesis in the prostate. In this experiment, the soybean isoflavone mixture consisting of 74% genistein and 21% daidzein was fed to F344 male rats which received DMAB s.c. injections and were implanted with silicon tubes containing TP throughout the experiment. Rats administered isoflavone mixture far less developed adenocarcinomas in the prostate and seminal vesicles than those maintained on control diet [58]. Genistein has also been reported to be effective in DMBA-induced rat ovarian carcinogenesis. Dietary administration of genistein caused significant reduction in the incidence of DMBA-induced ovarian adenocarcinoma in female Sprague-Dawley rats: 86% reduction by feeding of 25 ppm genistein and 100% reduction by feeding of 250 ppm genistein for 50 weeks [68]. In these animal models of carcinogenesis, circulating hormone levels and counts of oocyte and follicles were not different from controls, indicating that genistein inhibited the development of DMBA-induced carcinogenesis without showing toxicity to the endocrine or reproductive systems of mice [69]. In ovariectomized rats with DMBA exposure, a high incidence of uterine dysplasia was shown to be associated with E2 feeding, whereas the incidence was low in rats fed genistein [4].
Anticancer Activities of Genistein on Cancer Progression
After the normal cells are transformed into cancer cells in the process of carcinogenesis, they tend to growth abnormally and have the potential to invade or spread to other parts of the body [60]. In this article, the processes such as cancerous growth in an original place and invasion or metastasis are defined as cancer progression. Anticancer effects of genistein on the prevention of cancer progression separating its effects on the inhibition of carcinogenesis will be described in this section.
Firstly, the appropriate animal cancer models which provide the process of cancer progression are needed to evaluate the chemopreventive effect of genistein. Several transgenic animal models have been used for this purpose. Originally, transgenesis means the process of introducing an exogenous gene called a transgene into a living organism, which will exhibit a new property and transmit that property to its offspring [14]. In this case, transgenic animals are genetically modified to form certain types of cancer [10]. The transgenic models are useful tools in cancer research because it can be possible that cancer arises spontaneously in its natural tissue environment and progresses through multiple stages [76]. As a transgenic mammary cancer model, the mouse mammary tumor virus (MMTV)-neu/ErbB-2 transgenic mouse was developed to form spontaneous mammary cancer with a long latency due to overexpression of the neu proto-oncogene [26]. neu/ErbB-2 is equivalent to HER2 in humans, which is a member of the HER family of receptors and an important regulator of normal mammary growth and development. Overexpression of HER2 has been shown to play an important role in the development and progression of breast cancer [83]. Therefore, the MMTV-neu/ErbB-2 transgenic mouse model can provide the information about the influence of genistein on the genetic mutations associated with carcinogenesis in mammary cancer. When the MMTV-neu mice were fed AIN-93G diets containing 250 mg/kg genistein from 7 wk of age, mammary tumor latency was significantly delayed compared with a control. However, genistein did not reduce the number or size of tumors once tumors formed. Hence, this study showed that genistein has an effect of delaying the initiation of mammary cancer growth [31]. Specifically for prostate cancer research, the TRAMP model, an autochthonous transgenic animal model of prostate cancer, has been developed as a very important tool for understanding the earlier stage events and the progression of adenocarcinomas [24]. Because TRAMP mice can develop in situ and invasive carcinoma of the prostate, the whole spectrum of human prostate cancer progression from prostatic intra-epithelial neoplasia (PIN) to large multinodular malignant neoplasia may be realized from adopting this model [23,33]. Genistein also showed the promising chemopreventive effect against prostate cancer in the TRAMP models. When the treatments were 250 mg genistein/kg AIN-76A diet prepubertally only, in adulthood only or throughout life, poorly-differentiated (PD) cancerous lesions resulted in 6, 29, and 50% decreases compared with controls, respectively [76]. Other study showed that genistein in the diet of TRAMP/FVB mice induced a reduction in prostate size and the incidence of PD cancer ensuing in an accumulation of prostates at the PIN stage [16]. The underlying mechanism of genistein on the suppression of TRAMP prostate cancer progression was elucidated that it inhibited the activation of Akt, restored the activation of glycogen synthase kinase 3-beta (GSK-3β), and reduced cyclin D1 levels [16]. Dietary genistein improved the survival of TRAMP mice by reducing the expression of osteopontin in the prostate [47]. Osteopontin is an extracellular matrix protein secreted by macrophages infiltrating prostate tumors and may have a role in the transition from clinically insignificant tumors to metastatic prostate cancer [57,82].
Next, the mouse cancer models xenografted with diverse types of human cancer cell lines have been used for the evaluation of anticancer effects of genistein. In general, the xenografted mouse is manufactured by implanting cancer cell lines originally derived from patient tumors into an immune-compromised mouse [6,62]. As immune-compromised rodents, athymic nude mice and severely compromised immunodeficient (SCID) mice are commonly used in cancer research [52]. In addition, for the formation of hormone-responsive cancer models such as breast, ovarian, and prostate cancers, the ovariectomized or castrated mice with low circulating estrogen or androgen levels are used to exclude the hormone induced stimulation effects of these cancers or mimic the endogeneous estrogen state of postmenopausal women. Because anticancer agents or treatments are given to the mouse when the tumor volume reaches a certain size, this model is appropriate to evaluate their chemopreventive effects on cancer progression. According to the study using severe combined immunodeficient mice implanted with MCF-7 breast cancer cells, the diet supplemented with 0.07% genistein was found to significantly inhibit tumor growth [45]. In the study of female athymic nude mice xenografted with MDA-MB-435 breast cancer cells, the diet supplemented with 750 µg/g genistein reduced the tumor volume and inhibited metastatic burden in the lungs and lymph node metastasis [74]. A recent study demonstrated that genistein inhibits breast cancer stem-like cells (BCSCs) that are profound implications for cancer prevention [32] in nude mice xenograft models [18]. Also for ovarian cancer models, genistein has been reported to inhibit ovarian cancer progression by pleiotropic mechanisms [39]. In our previous study, genistein was determined to have an suppressing effect on tumor growth by inducing apoptotic signaling cascades in BG-1 ovarian cancer cell xenografted mice [29]. It was additionally found in this study that genistein negated the ovarian cancer proliferation induced by estrogen or bisphenol A (BPA), a typical endocrine disrupting chemical. Genistein significantly inhibited the proliferation of HO-8910PM ovarian cancer cell xenografted tumors by regulating cell cycle and apoptoic genes in the nude mice [77]. Moreover, in the endometrial cancer models, genistein (1 mg/30 g b.w./every 4 weeks for 30 weeks) suppressed the cancer development and atypical endometrial hyperplasia by regulating the expression of estrogen-related genes and cytokines [42]. Genistein has been also linked to lower mortality of prostate cancer in prostate cancer xenograft mouse models. In the athymic xenografted mice orthotopically implanted with human PC3-M prostate cancer cells, genistein decreased metastasis by 96% in contrast to the formation of lung metastasis in >80% of the mice [36]. Other study also showed that prostate cancer treatment was enhanced by genistein in the orthotopic xenograft nude mice implanted with human PC-3 prostate cancer cells [78]. Other studies also showed that genistein has an antitumor effect against other cancers. Genistein was also shown to be chemopreventive against bladder cancer growth and metastasis. In SCID mouse tumor model orthotopically implanted with 253J B-V human bladder cancer cells, genistein had reduced final tumor weights by 56% by induction of tumor cell apoptosis and inhibition of tumor angiogenesis in vivo [64]. In addition, in the mice transplanted with B16F-10 melanoma and Lewis lung cancer cells, genistein inhibited tumor growth in case of intradermal or subcutaneous injection of tumor cells and metastasis in case of intravenous or subcutaneous injection of tumor cells [80]. Genistein particularly showed antimetastatic effects by inhibiting lung metastasis induced by B16F-10 melanoma cells in C57BL/6 mice (53.6%) and by increasing the life span of the tumor-bearing animals (47.7%) [80].
Conclusion
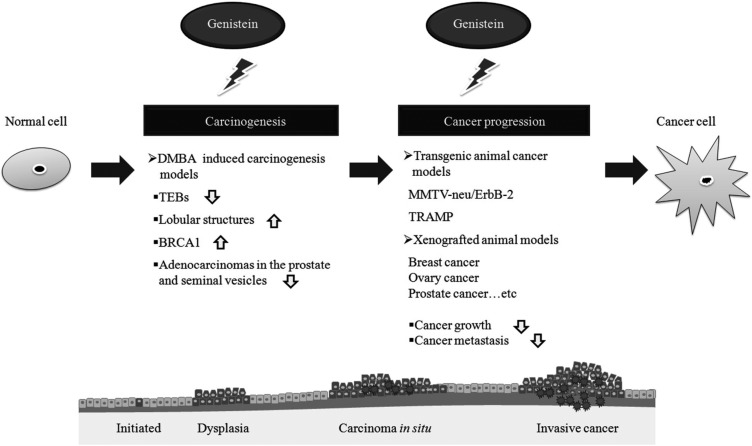
The present study indicates that genistein clearly exerts a chemopreventive effect against certain types of cancer. In particular, this study focused on the anticancer efficacy of genistein elucidated in the animal experiments in respect of a developmental stage of cancer; carcinogenesis in the earlier stage as well as cancer progression thereafter. It was collectively found that genistein effectively inhibits carcinogenesis by reducing the incidence of carcinogen-induced adenocarcinoma and cancer progression by suppressing the tumor growth and metastatic transition as shown in Figure 2. In spite of numerous outcomes and evidence supporting excellent anticancer efficacies of genistein, its protective effect has been controversial. The some previous studies insisted that genistein did not have any inhibitory effects on mammary carcinogenesis in female rats under estrogen-free conditions [71] and showed no significant reduction mammary tumor multiplicity in female SD rats [11]. Furthermore, genistein was reported to promote disease progression of prostate cancer in vivo [55] and stimulate tumor progression in advanced human prostate cancer by increasing epidermal growth factor receptor signaling [54]. Because genistein distinctively has a biphasic effect on its concentration or dosage, it seems to be possible that genistein may act as an agonist or an antagonist for a certain type of cancer. Nevertheless, the studies proving the chemopreventive potential of genistein have continued and the adjuvant roles of genistein in the conventional cancer therapy have been magnified. Genistein was found to enhance the efficacy of cabazitaxel chemotherapy in metastatic castration-resistant prostate cancer cells [84] and enhance radiotherapy in a metastatic prostate cancer model [61]. Taken together, it can be concluded that genistein may be a candidate for chemoprevention of carcinogenesis and cancer progression and may deserve to be the central compound supporting the epidemiological evidence stated above. More careful examination of the effects of genistein in a chemotherapeutic setting is needed [17] and further clinical studies have to be examined to establish the possibility of genistein as a chemopreventive agent.
 XML Download
XML Download