

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Group 2 innate lymphoid cells (ILC2s) were originally described as a novel population of lineage-negative cells that induced innate type 2 responses by producing the critical Th2-type cytokines IL-5 and IL-13 in response to IL-25 and IL-33.123 ILC2s are reported to contribute to helminth expulsion, airway hyper-responsiveness, airway remodeling, and homeostasis,24567 and also appear to serve as key players in the development of allergic inflammation. Indeed, house dust mite (HDM)-induced asthma is accompanied by increased numbers of ILC2s in lung and bronchoalveolar lavage fluid,8 and in murine models of asthma pulmonary ILC2s have been reported to produce over half of the IL-5 and IL-13 after intranasal administration of IL-25/IL-33.8 Similarly, human ILC2s have been found to be enhanced in the peripheral blood of patients with asthma and up-regulate innate type 2 immune responses via IL-25/IL-33-stimulated production of IL-5 and IL-13,9 thus highlighting the potentially critical role of ILC2s in allergic disease.
The relationship between allergic rhinitis (AR) and asthma is well established by the concept of a "united airway."10 AR is an inflammatory disease of the nasal mucous membrane, resulting as a consequence of allergen-mediated response of mast cells characterized by the release of a variety of Th2 cytokines, which have clear-cut effects on eosinophil survival and allergen-specific IgE synthesis.1112 More recently, Doherty et al.13 have reported that nasal challenge using cat allergen in cat-sensitized adults with a current history of rhinitis symptoms on exposure to cat significantly increase the percentage of ILC2s in the peripheral blood of these subjects, compared to baseline levels and diluent challenge, 4 hours after challenge. However, another recent study has demonstrated that the prevalence of ILC2s is greater in blood from subjects with allergic asthma than from those with AR and that PBMCs from asthmatics produce significantly greater amounts of IL-5 and IL-13 in response to IL-25 or IL-33 than those from AR subjects.9 The same study suggested that the differences in ILC2s and innate type 2 responses in these diseases may be a consequence of immunopathogenic differences. We hypothesized that the difference in the number of ILC2s may be related to the distinct clinical features of AR and/or to different allergen exposure. Thus, the aim of this study was to characterize the ILC2 population and to investigate the potential role of ILC2 in the distinct subtypes of AR monosensitized to HDM or mugwort.
MATERIALS AND METHODS
Study design and subjects
Nine AR patients monosensitized to Dermatophagoides pteronyssinus (D. pteronyssinus)/D. farinae (HDM-AR) from June to July in 2014 and 8 AR patients monosensitized to Artemisia argyi (mugwort-AR) in September 2014, the peak mugwort season, were recruited from the AR clinic at Beijing TongRen Hospital. Seven volunteers with no symptoms of AR and a negative skin prick test result to common allergens (D. pteronyssinus/D. farinae, animal hair, tree mix, grass mix, cereal mix, mugwort, dandelion, giant ragweed, Chenopodium album, Humulus species, locust bean, Blatellagermanica, pine, plantain, Curvularialunata, Candida albicans, Penicilliumnotatum, Alternaria tenuis, and Aspergillusfumigatus) were enrolled as healthy controls (HCs).
All participants rated the severity of their symptoms of rhinorrhea, sneezing, nasal obstruction, nasal pruritus, and ocular pruritus on a scale of 0-3 (0=symptom not present; 1=mild, symptom present but not bothersome; 2=moderate, symptom bothersome but easily tolerated; and 3=severe, symptom difficult to tolerate), which were expressed as the Total 5 Symptom Score (T5SS). All patients were required to have persistent AR according to the ARIA criteria,14 with 1 or more severe symptoms, as scored according to the T5SS scale, and equal or greater than class 2 serum-specific IgE against D. pteronyssinus/D. farinae or Artemisia argyi (EUROBlotMaster 44, Lübeck, Schleswing-Holstein, Germany) to be included in the study. Patient characteristics are summarized in Table. Patients who took antihistamines, steroids, or leukotriene receptor antagonists within 4 weeks, immunotherapy for any allergen, or had an allergic reaction to any drug within the last 2 weeks were excluded from the study. Similarly, patients who had asthma and those who had an acute infection within the last 4 weeks or were smokers within the past 12 months were excluded, as were pregnant patients. The study protocol was approved by the local ethics committee, and informed consent was obtained from each participant prior to enrolment in the study.
Analysis of ILC2s by flow cytometry
We followed the strategy used by Mjösberg et al.15 Whole blood cells were stained with a fluoresceinisothiocyanate (FITC)-conjugated lineage cocktail, phycoerythrin-CY7-conjugated anti-CD127 Ab and phycoerythrin-conjugated anti-CRTH2 Ab, or the respective isotype Abs (all from BD Pharmingen, San Diego, CA, USA), and a cell lineage cocktail comprising antibodies to CD3, CD14, CD16, CD19, CD34, CD123, CD11c, TCRαβ, and TCRγδ expressed on T cells, monocytes, macrophages, B cells, mast cells, dendritic cells, and hematopoietic progenitor cells. We gated on cells lacking lineage markers and examined expression of CD127 and CRTH2 within this population; with all Lineage-CD127+CRTH2+ lymphocytes considered ILC2s (Fig. 1). Finally, cell count was performed using a FACSAriaII flow cytometer (BD Biosciences, San Diego, CA, USA). Data were analyzed using FACSDiva software (BD Biosciences). The number of ILC2s is expressed as a percentage of all lymphocytes.
Peripheral blood mononuclear cells (PBMCs), and lineage-positive and-negative cells in response to IL-33 and IL-25 in vitro
Heparinized blood (10 mL) was obtained from all HMD-AR and mugwort-AR patients, and PBMCs were collected from the blood by Ficoll-Plaque Plus density gradient centrifugation. The PBMCs were stained with the FITC-lineage cocktail as described above and were separated into 2 fractions of Lineage+ and Lineage- cells based on the lineage markers, using a fluorescence-activated cell sorter (BD FACSAriaII; BD Biosciences). The Lineage+ and Lineage- cells were washed by suspension and centrifugation at 300 g for 8 minutes in fresh RPMI 1640 medium, and were then resuspended in RPMI 1640 media containing 10% fetal bovine serum (FBS), penicillin (100 U/mL), and streptomycin (100 mg/mL). PBMCs (1.5-2×106 cells/mL), Lineage+ cells, and Lineage- cells (2-2.5×105 cells/mL) were cultured in 96-well round-bottom plates at 37℃ with 5% CO2 in air atmosphere for 24 hours in the presence of 50 ng/mL IL-25, 50 ng/mL IL-33, or their combination (all from R&D Systems Inc., Minneapolis, MN, USA), together with IL-2 (20 U/mL), a cytokine necessary for lymphoid cell activation.16 At the end of culture, the cell-free supernatants were collected from each well and assessed for the presence of IL-5 and IL-13, using a Milliplex magnetic bead kit (EMD Millipore Corp, Billerica, MA, USA) as recommended by the manufacturer.
Statistical analysis
Qualitative variables were compared between the groups by χ2 tests, and continuous variables were analyzed by the nonparametric Mann-Whitney U test for unpaired comparison. Statistical analysis was performed using SPSS version 19.0 statistical software (IBM, Armonk, NY, USA) and graphs were generated using the prism software (version 4.00; GraphPad, La Jolla, CA, USA). All tests were 2-tailed, and P values of less than 0.05 were considered significant.
RESULTS
Clinical characteristics of the patients
A total of 17 AR patients monosensitized to HDM or mugwort and 7 HCs were enrolled in this study. Mugwort-AR patients were closely confined to the grass pollen season with allergic symptoms. There were no significant age, gender, or disease differences between the groups. Similarly, there were no significant differences in T5SS and specific IgE levels between the HDM-AR and mugwort-AR patient groups (Table).
Assessment of IL-5 and IL-13 released from the PBMCs of HDM/mugwort-AR patients
In the presence of IL-2, the release of IL-5 by PBMCs from AR patients was significantly increased by incubation with IL-33 alone and in combination with IL-25 compared to incubation with IL-25 alone (Fig. 3A; P=0.020 and 0.003, respectively), IL-2 alone (Fig. 3A; P=0.006 and 0.011, respectively), or medium (Fig. 3A; P=0.002 and 0.002, respectively). In contrast, IL-25 plus IL-2 did not significantly alter the levels of IL-5 compared to IL-2 alone or medium (Fig. 3A).
Similarly, in the presence of IL-2, the release of IL-13 from the PBMCs of AR patients was significantly increased by incubation with IL-33 alone and in combination with IL-25 compared to incubation with IL-25 alone (Fig. 3B; P=0.005 and 0.000, respectively), IL-2 alone (Fig. 3B; P=0.002 and 0.002, respectively), or medium (Fig. 3B; P=0.000 and 0.000, respectively). In addition, IL-25 plus IL-2 significantly induced higher IL-13 production from PBMCs compared to IL-2 alone (Fig. 3B; P= 0.034), or medium (Fig. 3B; P=0.008).
Subgroup analysis between HDM-AR and mugwort-AR patients further demonstrated that IL-33 plus IL-2 as well as the combination of IL-33 and IL-25 plus IL-2 induced a significantly greater release of IL-5 (Fig. 4A; P=0.048, 0.014, respectively) and IL-13 (Fig. 4B; P=0.011 and 0.011, respectively) from the PBMCs of HDM-AR patients compared with mugwort-AR patients. IL-13 release was also significantly higher in the PBMCs of HDM-AR patients stimulated with IL-25 plus IL-2 compared to mugwort-AR patients (Fig. 4B; P=0.010).
IL-5 and IL-13 released from Lineage cells
Since an extremely small number of ILC2s were present in PBMCs (Fig. 2), ILCs were not suitably obtained for further culture experiments. Assessment of IL-33- and/or IL-25-induced release of IL-5 and IL-13 from the total Lineage-positive and total linage-negative cells in the presence of IL-2 demonstrated that Lineage- cells released significantly greater amounts of IL-5 compared to Lineage+ cells following stimulation with IL-33+IL-25+IL-2, IL-33+IL-2, and IL-25+IL-2 (Fig. 5A; P=0.021, 0.020, and 0.021, respectively). Similarly, Lineage- cells also released significantly greater amounts of IL-13 compared to Lineage+ cells after stimulation with IL-33+IL-25+IL-2, IL-33+IL-2, and IL-25+IL-2 (Fig. 5B; P=0.020, 0.021, and 0.043, respectively).
DISCUSSION
This study demonstrated that Chinese HDM-AR patients contained a significantly higher level of blood ILC2s compared to mugwort-AR patients and HC subjects. Furthermore, both IL-5 and IL-13 were released in significantly greater amounts from the PBMCs of HDM-AR patients in response to IL-33 and/or IL-25 combined with IL-2 than from similarly stimulated PBMCs from mugwort-AR patients; and Lineage- cells, rather than Th2 cells, were the main source of IL-5 and IL-13 in the HDM-AR patients. These findings suggest that ILC2s induce major innate type 2 immunity and play a predominant role in the development of HDM-AR.
Our finding that the percentage of ILC2s was not increased in the group of patients with mugwort-AR is in accordance with those of a recent study by Bartemes et al.9 However, another recent study by Lao-Araya et al.17 has demonstrated that the proportion of ILC2s was elevated in seasonal Timothy grass (Phleum pratense)-sensitized AR subjects, 66.7% of whom were polysensitized to HDM allergen. This finding suggests that HDM allergens per se may possibly show higher immunogenicity toward ILC2s than plant allergens, such as mugwort pollen, or may stimulate inflammatory responses through different mechanisms from pollen allergens, which lead to differential generation of ILC2s. Indeed, while allergic immune responses to the major mugwort pollen allergen Art v 1, a modular glycosylated protein,18 are characterized by IgE binding and T-cell proliferation;1819 HDM-mediated direct nonspecific damage and allergic reactions in the respiratory epithelium2021 are induced by trypsin/chymotrypsin-like enzymatic activities associated with mite allergens.2223 These findings support the concept that sensitization to HDM allergens may potentially lead to greater epithelial cell-derived IL-33 release and generation of systemic ILC2s than sensitization to mugwort allergen, and thus provide a plausible explanation for the clear differences in ILC2s numbers noted between HDM- and mugwort-allergic patients in our study.
In this study, IL-33 in the presence of IL-2 was a strong stimulator of ILC2s compared to IL-25 in the presence of IL-2, and ILC2s induced acute innate inflammation 24 hours after culture with IL-33 and IL-2, even after 5 days of chronic stimulation.9 IL-33, an epithelial cell-derived cytokine, has been shown to be significantly increased in the nasal secretions of subjects with AR compared to the nasal secretions of healthy control subjects,24 and play an important role in the pathogenesis of allergic sensitization.25 Furthermore, IL-33 has been shown to be a potent stimulus for ILC2s activation and migration in vitro,2627 and promote the expansion of ILC2s into the airway in the initiation of HDM-induced Th2 immunity.25 Collectively, these results raise the possibility that recruitment and activation of ILC2s in AR could be triggered by humoral factors, such as IL-33, which are released from the nasal mucosa of AR following exposure to allergens.
Although no relationship between circulating ILC2s and symptom scores of AR patients was found in this study (data not shown), it is possible that such a relationship may be dependent on a correlation between the number of ILC2s in the nasal mucosa of AR patients and the severity of disease in these individuals. Several studies have demonstrated that ILC2s are enriched in patients with nasal polyps and are positively associated with Th2-cell proportion and eosinophil count.28293031 Moreover, endoscopic sinus surgeries or systemic corticosteroid treatment have been shown to be correlated with reduced polyp ILC2s,2930 suggesting that a decrease in ILC2s at the sinonasal mucosal barrier may relieve ongoing local inflammation. Clearly, further research into the precise roles of ILC2s in the nasal mucosa and the regulatory pathway influencing the expression of epithelial and Th2 cytokine in the context of AR is required, along with a better understanding of the allergic onset, exacerbation, or progression of the ILC2s-mediated inflammation. Thus, targeting ILC2s could be a potential therapeutic approach in the treatment of AR, although this needs to be investigated in large well-designed trials.
In summary, this study demonstrated that the relative number of ILC2s in the total lymphocyte fraction was significantly increased in the blood of Chinese subjects with HDM-AR, but not in subjects with mugwort-AR. It is possible that differences in ILC2s frequencies between the 2 groups of patients may be related to differences in the allergenicity of sensitizing agents. Importantly, however, ILC2s may play an important role in the pathogenesis of HDM-AR by producing large amounts of IL-5 and IL-13 in response to IL-25 and/or IL-33.


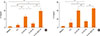



 XML Download
XML Download