

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Rice (Oryza sativa L.) is widely cultivated in the world, and half of the world's population depends on rice as their main source of food [1]. The outer bran and germ portion of intact rice grains (brown rice) has gained popularity because of its high contents of multiple nutrients, including fiber, vitamins, minerals, and phytochemicals [2]. Consumption of brown rice has been shown to decrease serum cholesterol concentration in human [3,4] and hypercholesterolemia-induced animals [5,6,7], suggesting that polyphenols in brown rice may have a hypocholesterolemic effect [2].
Pigmented rice bran, such as black, red, and purple rice has also been reported to have higher health promoting potential depending on the type, contents, and structural properties of phenolic compounds [8,9,10]. Most studies focused on the anti-oxidative effect of phenolic compounds [11,12,13,14]; however, those have also been known to reduce serum concentration of cholesterol in vivo [15,16,17], possibly through inhibition of 3-hydroxyl-3-methylglutaryl coenzyme A (HMG-CoA) reductase and acyl-coenzyme A cholesterol acyltransferase-2 (ACAT-2).
Consumption of black rice [11,18] and black rice extract [12,19] has been shown to reduce serum cholesterol concentration in dyslipidemic animals. Red rice intake also reduced the ratio of LDL/HDL-cholesterol in rabbits fed high cholesterol diets [11], however, little is known about regulation of cholesterol metabolism by bioactive compounds in pigmented rice.
We previously reported that the developed red pericarp glutinous rice (red rice), Jakwangchalbyeo, contained seven times more total phenolic compounds, mainly ferulic acid, hesperidin, and catechin, than brown rice [20]. Therefore, the current study investigated the hypothesis that the newly developed red rice rich in polyphenols has cholesterol lowering effects through regulation of protein expression linked to synthesis and degradation of cholesterol in hypercholesterolemic mice as compared with brown rice.
MATERIALS AND METHODS
Animals and diets
The protocol was approved by the Institutional Animal Care and Use Committee of Hanyang University (HY-IACUC-12-026). Five-week old C57BL/6 male mice (Orient Bio, Gyeonggi-do, Korea) were housed in individual ventilated cages in an airconditioned room maintained at 22 ± 2℃ with a 12 h light-dark cycle. After acclimatization for one week, 20 mice were randomly divided into four isoenergetic diet groups (Table 1; n = 5 each); American Institute of Nutrition (AIN)-93G diet (normal control, NC), AIN-93G diet with 2% cholesterol (high cholesterol, HC), brown rice with 2% cholesterol (BR-HC), or red rice with 2% cholesterol (RR-HC). Amount of cholesterol in the diet was previously confirmed to cause development of hypercholesterolemia in a rodent model [21,22,23,24].
Jakwangchalbyeo (red pericarp glutinous rice; red rice) and Dongjinchalbyeo (white pericarp glutinous rice; brown rice) were cultivated at the Konkuk University Experimental Farm. Dongjinchalbyeo is widely cultivated and consumed in Korea. Harvested rice seeds were dried, hulled using a milling machine, and ground by a pin-type mill (DK-201, Sejung Tech, Daegu, Korea). We previously reported that red rice contained 1245 µg/g of phenolic compound, such as 83.4 µg/g of ferulic acid, 39.8 µg/g of catechin, and 28.0 µg/g of hesperidin, while brown rice contained 159 µg/g of phenolic compound [20].
Diet and water were allowed ad libitum for 12 weeks. Food intake was measured daily and body weight was measured weekly. At the end of the experiment, mice were fasted overnight and euthanized by exsanguination under anesthesia with an intraperitoneal injection of tiletamine (25 mg/kg), zolazepam (25 mg/kg), and xylazine (10 mg/kg). Blood was collected into serum separation tubes (BD Vacutainer, Franklin Lakes, NJ, USA) from the abdominal aorta and centrifuged at 3,000 g for 15 min (HA 1000-3, Hanil Sciences Industrial CO. Ltd., Incheon, Korea). Organs were harvested, rinsed with saline, and then weighed. Serum and tissue samples were stored at -80℃ until further analysis.
Biochemical assays
The serum levels of triglyceride, total-cholesterol, low-density lipoprotein (LDL)-cholesterol, alanine transaminase (ALT), and aspartate transaminase (AST) were determined using commercial kits (IDLabs Inc., London, Ontario, Canada). All experiments were performed according to the manufacturer's instructions and the results were measured using a microplate reader (iMark Microplate Reader, BioRad, Richmond, CA, USA).
Western blotting
Liver tissues were homogenized in a 0.8 mL ice-cold lysis buffer (0.25 M sucrose, 20 mM HEPES, 2 mM dithiothreitol, 0.5 mM EDTA, 10 µg/mL leupeptin, 1 mM PMSF, 10 µg/mL aprotinin, and 1 mM Na3VO4, pH 7.5). The homogenous liver tissues were centrifuged at 10,000 g for 15 min at 4℃ (Eppendorf, Hamburg, Germany), and the supernatant was centrifuged at 20,000 g for 1 h at 4℃ to obtain the cytosolic fraction. The protein concentrations were determined using a Bradford assay with bovine serum albumin (Bio-Rad, Hercules, CA, USA) as the standard. Equal amounts of protein (30 µg) were separated on 8% SDS-PAGE and transferred to a polyvinylidine fluoride membrane (0.45 µm, Immobilon-P transfer membrane, Millipore, Billerica, MA, USA). Membranes were blocked for 1 h at room temperature with 5% skim milk in Tris-buffered saline with Tween 20 (TBST) and then incubated with a primary antibody for ACAT-2 (1:250, Cayman chemical, Ann Arbor, MI, USA), sterol regulatory element binding protein-2 (SREBP-2, 1:500, abCam, Cambridge, UK), HMG-CoA reductase (1:100, Santa Cruz Biotechnology, Santa Cruz, CA, USA), adenosine monophosphate activated protein kinase (AMPK, 1:1,000, Cell Signaling Technology, Beverly, MA, USA), phosphorylated AMPK (p-AMPK, 1:500, Cell Signaling Technology, Beverly, MA, USA), cholesterol 7-α-hydroxylase (CYP7a1, 1:1,000, abCam, Cambridge, UK) or sterol 12-α-hydroxylase (CYP8b1, 1:100, Santa Cruz Biotechnology, Santa Cruz, CA, USA) in TBST containing 5% skim milk at 4℃. After washing three times with TBST, the membranes were incubated with goat anti-rabbit (1:20,000, Santa Cruz Biotechnology, Santa Cruz, CA, USA) or mouse IgG secondary antibody (1:5,000, Cell Signaling Technology, Beverly, MA, USA) for 1 h at room temperature. The immunoreactive bands were developed using the enhanced chemiluminescence kit (GE Healthcare Life Sciences, Piscataway, NJ, USA) and exposed to X-ray film (Kodak Inc., Rochester, NY, USA). The relative amounts of protein were calculated by normalizing the value to β-actin (1:1,000, BD Transduction Laboratories, NJ, USA).
Liver lipid extraction
The lipid extract with Folch solution was centrifuged at 3,000 g for 10 min to obtain the supernatants. Hepatic triglyceride and total cholesterol were determined using a commercial kit (Asan Pharm., Hwaseong, Korea) with a spectrophotometer (DU 600, Beckman Coulter, Inc., Indianapolis, IN, USA).
Immunofluorescence
Liver (~4 mm) was fixed in 4% paraformaldehyde in phosphate-buffered saline at 4℃ for 24 h, and paraffin sections were cut and deparaffinized using ethanol and xylene. After incubation in 10% normal goat serum with 0.3% Triton X-100 for 1 h for blocking, samples were incubated with primary antibody (1:200, AbCam, Cambridge, UK) in 10% normal goat serum overnight at 4℃. After rinsing, secondary antibody (1:500, Invitrogen, Carlsbad, CA) was applied with 10% NGS at room temperature for 2 h in the dark. The nuclei were stained with 4, 6-Diamidino-2-phenylindole (DAPI) and the sections were mounted. The slides were left in the dark overnight and then sealed using clear nail polish. Images were then taken using a Leica microscope (Leica Microsystems GmbH, Wetzlar, Germany) and Leica Application Suite Advanced Fluorescence software (Leica Microsystems GmbH, Wetzlar, Germany).
Statistical analysis
Values were expressed as mean ± standard error of mean (SEM). Statistical differences among the groups were calculated by analysis of variance (ANOVA) followed by Duncan's multiple range test (SPSS 18.0 version., Chicago, IL, USA). Differences with P < 0.05 were considered significant.
RESULTS
Food intake, bodyweight, and organ weight
There were no significant differences in dietary intake, initial and final body weight, and weight of kidney, muscle, heart, visceral fat, and epididymal fat (Table 2). However, weight of liver was significantly higher in mice with HC, BR-HC, and RR-HC than in those with NC.
Serum and hepatic levels of lipids
Serum triglyceride concentrations did not differ significantly between groups, but serum concentrations of total cholesterol, alanine transaminase, and aspartate transaminase were significantly higher in HC, BR-HC, and RR-HC than in NC (Table 3). However, serum concentration of LDL-cholesterol, and hepatic concentration of triglyceride and total cholesterol were significantly lower in RR-HC and BR-HC than in HC, and similar to that in NC.
Expression of protein associated with cholesterol metabolism
Expression of ACAT-2 and SREBP-2 was significantly decreased, but the expression of p-AMPK/AMPK ratio was significantly increased in RR-HC, BR-HC, and NC compared with HC (Fig. 1). In particular, the effect of red rice on expression of SREBP-2 was greater than that of brown rice. Expression of HMG-CoA reductase was significantly increased in HC, but decreased in RR-HC as compared with NC and BR-HC.
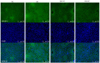
Immunofluorescence staining of liver also showed increased expression of SREBP-2 (light green) in HC compared with NC, BR-HC, and RR-HC (Fig. 2). In addition, more foam cells (black circle) were detected in HC than in NC, BR-HC, and RR-HC. DAPI (nucleus) showed no difference in all mice. As shown in Fig. 2, SREBP-2 expression in mice fed red rice was consistently more similar to that of normal control than that of mice fed brown rice.
Expression of CYP7a1 and CYP8b1 was significantly increased in RR-HC, BR-HC, and NC compared with HC (Fig. 3). In particular, CYP8b1 expression showed a greater increase in mice fed red rice than in mice fed brown rice, suggesting that the effect on cholesterol degradation was greater by red rice than brown rice.
DISCUSSION
The current study demonstrated that consumption of red rice rich in polyphenols caused a decrease in serum and hepatic level of LDL-cholesterol in mice fed a high cholesterol diet, which was due to inhibition of hepatic cholesterol synthesis by the decreased expression of HMG-CoA reductase, ACAT-2 and SREBP-2 expression, and the increased degradation of cholesterol by the increased expression of CYP7a1 and CYP8b1. Brown rice had a similar effect on cholesterol metabolism, but red rice showed greater effects, as we hypothesized.
Previous studies consistently showed that pigmented rice contained higher levels of phenolic compounds [8], and in particular, red rice contained more phenolic compounds than black rice [9]. Only a few studies of red rice have been reported; effect of red rice on radical-scavenging activity and antioxidant capacity [13]. Only one previous study examining the effect of red rice on cholesterol showed an increase in plasma HDL-cholesterol concentration and antioxidant status in rabbits fed a high cholesterol diet, but not in the concentration of LDL-cholesterol or total cholesterol [11]. Other pigmented rice, black rice and black rice extracts decreased serum cholesterol concentration in vivo [11,12,18,19], suggesting that cholesterol degradation and/or synthesis were modulated.
Salgado et al. [18] reported that the hypocholesterolemic effect of black rice was mediated by increasing cholesterol excretion in hypercholesterolemic rats. Similarly, brown rice intake has been shown to result in up-regulation of cholesterol catabolism by enhancing fecal bile acid excretion and improving the activity of CYP7a1 in hepatoma-bearing rats [6]. Consumption of rice protein also resulted in increased biliary secretion and fecal excretion of bile acid, and expression CYP7a1 in hypercholesterolemic rats [25]. Cholesterol 7-α-hydroxylase is the rate-limiting enzyme for cholesterol degradation to bile acid, and regulates CYP8b1 [26]. In the current study, expression of CYP7a1 and CYP8b1 was significantly increased in mice fed red and brown rice. In particular, CYP8b1 expression showed a greater increase in mice fed red rice than in mice fed brown rice, suggesting that the effect on cholesterol degradation was greater by red rice than brown rice.
Although the identities of the bioactive compounds in pigmented rice and the mechanisms involved are not well known, phenolic compounds such as ferulic acid [15,27], hesperidin [28], and catechin [16,17] have consistently been reported to decrease serum levels of total cholesterol and LDL-cholesterol in vivo. Catechin has also been shown to increase cholesterol degradation by activation of CYP7a1 in mice fed a high fat diet [17], and fecal cholesterol excretion in hamsters fed a high fat diet [29].
Phenolic compounds have also been shown to reduce hepatic cholesterol synthesis by regulating the expression of HMG-CoA reductase, the rate-limiting enzyme in the de novo synthesis of cholesterol [30,31,32]. In addition, HMG-CoA reductase is associated with SREBP-2, AMPK, and ACAT-2 in hepatic cholesterol synthesis [33,34].
Administration of ferulic acid and hesperidin resulted in decreased hepatic activity of HMG-CoA reductase and ACAT in apolipoprotein E-deficient mice fed a western diet [15] and rats fed a high cholesterol diet [16], respectively. Catechin also decreased cholesterol synthesis by inhibition of HMG-CoA reductase and SREBP-2 in mice fed a high fat diet [17], and by inhibition of HMG-CoA reductase and p-AMPK in hepatoma cells [30]. In addition, Kim et al. [33] suggested that individual phenolic compounds had a cholesterol lowering effect, and combination of polyphenol had a synergistic effect. Pigmented rice, particularly red rice contained several types of phenolic compounds [35], however, their effect on cholesterol metabolism has not been studied. Therefore, this was the first study to examine how consumption of pigmented rice can regulate cholesterol metabolism in vivo. In the current study, red rice consumption resulted in significantly decreased synthesis of hepatic cholesterol by modulating the expression of ACAT-2, SREBP-2, HMG-CoA reductase, and p-AMPK/AMPK ratio. In addition, immunofluorescence staining of liver consistently showed increased expression of SREBP-2 in mice fed red rice.
The current study had a limitation. Red rice and brown rice contain not only phenolic compounds but also dietary fiber, protein, fatty acids, vitamins, and minerals, which may also affect hepatic cholesterol metabolism. Thus, we were unable to determine which components of red rice were responsible for the hypocholesterolemic effect. However, red rice was more potent than brown rice, which could be due to the higher phenolic compounds in red rice.
In conclusion, red rice had a hypocholesterolemic effect by lowering hepatic cholesterol synthesis through HMG-CoA reductase, ACAT-2, p-AMPK/AMPK, and SREBP-2, and by enhancing hepatic cholesterol degradation through CYP7a1 and CYP8b1 in mice fed a hypercholesterolemic diet. Red rice may therefore have potential use in treatment and prevention of pathological states associated with cholesterol metabolism. Care may be required, but polyphenol rich red rice could be used in hypercholesterolemic patients, particularly those who consume a high carbohydrate diet.





 XML Download
XML Download