

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Sulfur is the seventh abundant mineral in body, and human weighing approximately 70 kg contains roughly 140 g of sulfur in body. Sulfur is consumed through food primarily in the form of sulfur-containing amino acids (SAAs), such as methionine, cysteine, cystine, and taurine, and in its glucosinolate form that is found in cruciferous vegetables, such as cabbage and cauliflower [1]. Sulfur is present in the sulfate form in water at a concentration of < 2 mg-> 1 g/L [1]. Moreover, sulfur can be ingested in inorganic forms, such as sulfite, sulfur dioxide, bisulfate, and met bisulfate, via preservatives that are added to processed food and drinks [2]. There is concern about toxicity of sulfur. Sulfide is produced when sulfur is incubated with the colon contents, and, as sulfide is associated with such symptoms as fatigue, drowsiness, and inhibited motor function [3]. However, toxicity due to inorganic sulfur is uncommon and appears to be restricted to the skin in the majority of cases [4]. When 0.17 mg/kg body weight of inorganic sulfur was orally administered to human, no toxicity was reported, nor was toxicity reported in animal tests using rabbits, rats, guinea pigs, and dogs [5]. The physiological effects of sulfur are mainly mediated by the organic form of sulfur. The anti-carcinogenic effects of organic sulfur were investigated via the sulfur-containing amino acids, (i.e., isothiocyanates, diallyl sulfide, allicin, glutathione, and α-lipoic acid) that are found in cruciferous plants, garlic, and methylsulfonylmethane (MSM), which is a byproduct of the wood pulp industry [6]. Sulfur is restrictively used in the treatment of dermatological disorders [3], psoriasis, rheumatic pain, and infections [7,8].
However, when inorganic sulfur is removed from the animal diet (even if there are sufficient SAAs for growth), avitaminosis E appears, collagen metabolism is damaged [9] and diet efficiency is reduced [10]. It is reported that pigs fed with high inorganic sulfur had an increase in SOCS3 mRNA in the ileum of pigs and thus likely counteracts the inflammatory activity of increased cytokine levels and minimizes cellular proliferation. High-S diet decreased the abundance of commensal bacteria with anti-inflammatory characteristics [11]. Inorganic sulfur is absorbed by active transport in the small intestines [12]. Inorganic sulfur also supplies the sulfur needed for cysteine formation and affects the functions of various physiological substances of which cysteine is a precursor [13].
Recently, it has been reported that the intake of refined inorganic sulfur reduces the clinical side effects of radiotherapy in cancer patients [14]. It was reported that when a daily dose of 0.5-1 g of refined sulfur was consumed in 24 cervical cancer patients, the sulfur, acting as a free radical scavenger, protected cells against DNA damage and reduced the side effects of radiotherapy. However, research on how inorganic sulfur can have anti-carcinogenic effects have not been well studied. Thus, we investigated the effect of inorganic sulfur on the inhibition of cell proliferation via regulation of epidermal growth factor receptor (EGFR) expression in MDA-MB-231 human breast cancer cells.
Materials and Methods
Reagents
Inorganic sulfur powder with a purity of 99% or more was obtained from Sulfon PS, Inc. (Seoul, Korea). The inorganic sulfur powder was dissolved in methyl alcohol (Sigma Aldrich, St. Louis, MO, USA) at a concentration of 5 mmol and stored at -20℃. Dulbecco's modified Eagle's medium/Ham's F12 Nutrient Mixture (DMEM/F12) along with streptomycin and penicillin was obtained from Gibco/BRL (Grand Island, NY, USA). Antibodies against ErbB2, ErbB3, pErbB2, pErbB3, Bcl2, Bax, and pAkt were purchased from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA, USA), and an antibody against Akt was purchased from Cell Signaling Technology, Inc. (Danvers, MA, USA). RIA-grade bovine serum albumin (BSA), transferrin, and other reagents were purchased from Sigma (St. Louis, MO, USA).
Cell culture
MDA-MB-231 human breast cancer cells were purchased from the American Type Culture Collection (Rockville, MD, USA). The cells were maintained in DMEM/F12 medium containing 100 mL/L of fetal bovine serum (FBS), 100,000 U/L of penicillin and 100 mg/L of streptomycin. The medium was replaced every 2-3 days. To examine of the effects of inorganic sulfur on breast cancer cell proliferation, the MDA-MB-231 cells were plated onto 24 well plates at a density of 2.5 × 104 cells/mL in DMEM/F12 medium supplemented with 10% FBS. After incubation for 48 h, the resulting cell monolayers were serum-starved with DMEM/F12 medium supplemented with 5 µg/mL of transferrin, 5 ng/mL of selenium, and 1 mg/mL of bovine serum albumin for 24 h. After serum starvation, the monolayers were incubated in serum-free medium (SFM) with 0, 12.5, 25, or 50 µmol/L of inorganic sulfur powder. The number of viable cells was estimated 24, 48, or 72 h after the cells were exposed to inorganic sulfur using the 3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide (MTT) assay as previously described [15]. The experiments were performed independently three times.
Western blot analysis
Cell lysates were prepared as previously described [16]. The total cell lysates were resolved on a 40-200 g/L sodium dodecyl sulfate polyacrylamide gel and were transferred to a polyvinylidene fluoride membrane (Millipore, Bedford, MA). Next, the blot was blocked for 1 h in 10 g/L BSA in TBS-T (20 mmol/L of Tris-Cl, 150 mmol/L of NaCl, 1 g/L of Tween-20, pH 7.5) or 50 g/L of milk TBS-T after which it was incubated for 1 h with the appropriate antibody (ErbB2, pErbB2, ErbB3, pErbB3, Akt, p-Akt, Bcl2, or Bax). Next, the blot was incubated with an HRP-conjugated anti-mouse or anti-rabbit secondary antibody. The signals produced during the assay were detected using the enhanced chemiluminescence method and the Super-Signal West Dura Extended Duration Substrate (Pierce, Rockford, IL). Finally, the relative abundance of each protein band was analyzed by scanning the exposed films densitometrically using the Image J Launcher (provided by NCBI).
Reverse transcriptase polymerase chain reaction
Total RNA was isolated using TRIzol reagent (Sigma), and cDNA was synthesized from 2 µg of total RNA using SuperScript II reverse transcriptase (lnvitrogen). To amplify the cDNA, primers for ErbB2 (upstream primer, 5'-CAAGAGTGCACGGCAGAGT-3'; downstream primer, 5'-GCCTTACAATGTGGGCATG-3'; annealed at 72℃ for 30 sec for 45 cycles), ErbB3 (upstream primer, 5'-CAAGAGTGCACGGCAGAGT-3'; downstream primer, 5'-GCCTTACAATGTGGGCATG-3'; annealed at 72℃ for 30 sec for 34 cycles), Akt (upstream primer, 5'-CAACTTCTCTGTGGCGCAGTG-3'; downstream primer, 5'-GACAGGTGGAAGAACAGCTCG-3'; annealed at 72℃ for 1 min for 30 cycles), Bcl2 (upstream primer, 5'-TGTGGATGACTGAGTACCTGAAC-3'; downstream primer, 5'-AGCTTTGTTTCATGGAACATCACTGAC-3'; annealed at 72℃ for 90 sec for 30 cycles), Bax (upstream primer, 5'-ATGGAGGGGTCCGGGGAG-3'; downstream primer, 5'-TGGAAGAAGATGGGCTGA-3'; annealed at 72℃ for 40 sec for 40 cycles) were used. The expression of human β-actin transcripts was examined as an internal control as described previously [17]. The PCR products were separated on a 1% agarose gel and were stained with ethidium bromide. The bands corresponding to each specific PCR product were quantified after the densitometric scanning of the exposed film using the Bio-Profile Bio-IL application (Vilber-Lourmat).
Statistical analysis
Statistical analyses were performed using the Statistical Analysis System software (SAS Institute, Cary, NC, USA). The data were expressed as means with their associated standard errors and analyzed with an analysis of variance (ANOVA). Any statistically significant differences among the means of the groups were tested at α = 0.05 using Duncan's multiple range test.
Results
Inorganic sulfur inhibits cell proliferation in MDA-MB-231 cells
A MTT assay was performed to examine the effect of inorganic sulfur on the proliferation of cancer cells. Treatment with inorganic sulfur for up to 24 h did not affect cancer cell proliferation, but treatment for 72 h significantly decreased cell proliferation in a dose-dependent manner (P < 0.05) (Fig. 1).
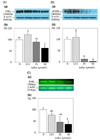
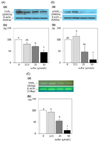
Inorganic sulfur reduces the protein and mrna expressions of ErbB2, ErbB3, and akt in MDA-MB-231 cells
The protein expression of ErbB2 and its active form, pErbB2, were significantly reduced following treatment with inorganic sulfur at concentrations of 25 µmol/L and 50 µmol/L, respectively (P < 0.05) (Fig. 2-A, 2-B). The protein expression of ErbB2 with the treatment of an inorganic sulfur concentration of 50 µmol/L was fell to 63.3% of the untreated control group. The protein expression of pErbB2 at the concentrations of 25 µmol/L and 50 µmol/L of inorganic sulfur was decreased to 13.1% and 2.4% of the untreated control group following treatment with, respectively. The mRNA expression of ErbB2 at a concentration of 50 µmol/L inorganic sulfur was significantly decreased to 37.2% (P < 0.05) of the level of the untreated control group (Fig. 2-C). The protein expression of ErbB3 significantly was decreased to 71.3% and 45.2% of the untreated control group following treatment with concentrations of 25 µmol/L and 50 µmol/L of inorganic sulfur, respectively (P < 0.05) (Fig. 3-A). The active form, pErbB3, was also significantly decreased to 47.2% and 30.3% of the untreated control group following treatment with concentrations of 25 µmol/L and 50 µmol/L of inorganic sulfur, respectively (P < 0.05) (Fig. 3-B). The mRNA expression of ErbB3 with the treatment of inorganic sulfur concentration of 25 µmol/L was also significantly decreased (P < 0.05) (Fig. 3-C). The protein and mRNA expressions of Akt were significantly decreased (P < 0.05) to 84.5% with the addition of 25 µmol/L inorganic sulfur and 62.4% of the level with the addition of 50 µmol/L, compared with untreated control group (P < 0.05) (Fig. 4-A, 4-C). However, the protein expression of pAkt was shown to be unaffected by treatment with inorganic sulfur (Fig. 4-B).
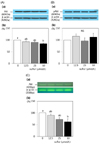
Inorganic sulfur increase the protein and mRNA expression of bax, but does not influence the protein and mRNA expression of Bcl2 in MDA-MB-231 cells
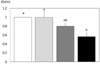
The protein and mRNA expression of Bax were significantly increased with the treatment of 50 µmol/L inorganic sulfur. The protein expression of Bax was significantly increased to 150.3% of the untreated control group following treatment with an inorganic sulfur concentration of 50 µmol/L (P < 0.05) (Fig. 5-A). The mRNA expression was also significantly increased to 398.7% of the untreated control group under the same conditions (P < 0.05) (Fig. 5-B). The protein and mRNA expression of Bcl2, a member of the Bcl family that inhibits cell apoptosis, were not affected by inorganic sulfur treatment (Fig. 6-A, 6-B). Consequently, the Bcl2/Bax protein expression ratio that is used as an indicator of apoptosis was significantly reduced to 56.4% of the ratio observed in the untreated control group (P < 0.05) following treatment with an inorganic sulfur concentration of 50 µmol/L (Fig. 7).
Discussion
This study investigated the effects of inorganic sulfur on cancer cell proliferation and the protein and mRNA expression of proteins that are related to cell proliferation and apoptosis in MDA-MB-231 cells. There is concern about toxicity of sulfur. Toxicity by inorganic sulfur is usually limited to the skin [4], there is no toxicity was reported from oral administration of 0.17 mg/kg inorganic sulfur to humans, or in other animals [5].
Diverse instances of cancer cell proliferation inhibition due to treatment with organic sulfur have been reported [18-20], but the inhibition of breast cancer cell proliferation by inorganic sulfur is reported here for the first time. When treated with inorganic sulfur, there is no effect on cell proliferation following an incubation of up to 24 h, but cell proliferation was inhibited in a dose-dependent manner after 72 h. Kong et al. [18] showed that the treatment of AGS human gastric adenocarcinoma cells with extracts from young radishes that had a high organic sulfur glucosinolate content showed the cancer cell growth inhibition. In addition, Bak et al. [19] reported that treating HT-29 human colon cancer cells with Kimchi extract that made with sulfur-treated radishes likewise resulted in cancer cell growth inhibition. Choi and Kim [20] reported that, when diverse cancer cells were treated with extracts from a hot-water extraction from regular ducks or organic sulfur-fed ducks, a noticeable effect in proliferation inhibition was seen in the cells treated with the organic sulfur-fed duck extract. L-sulforaphane, the organic sulfur compound that is abundant in cruciferous plants, induces apoptosis in HT-29 colon cancer cells [21] and in PC-3 human prostate cancer cells [22].
The epidermal growth factor receptor (EGFR) family consists of ErbB1, ErbB2, ErbB3, and ErbB4. When these members combine with an outside ligand, they interact to form a heterodimer that activates the intercellular kinase domain, and an external signal is triggered inside the cell as the tyrosine portion of the receptor's cytoplasmic domain undergoes autophosphorylation. Of the ErbB members, ErbB3 alone forms a heterodimer with ErbB2 to activate the tyrosine kinase [23-25]. ErbB2 plays an important role in breast cancer cell development in rats [26], and when ErbB2 expression is high in the normal vascular epithelia near the carcinoma, there is a strong likelihood that the tissue will transform into cancer cells [27]. Therefore, it has been reported that the inhibition of excessive EGFR expression is closely related to the inhibition of cancer cell proliferation [28,29]. EGFR delivers an external signal to the cell through the PI3K/Akt pathway [23]. Akt inhibits the activation of pro-caspase-9 by inducing the phosphorylation of pro-caspase-9, and by inducing Bax and the pro-apoptotic transcription factor FKHR to undergo phosphorylation [30-32]. AKT promotes growth factor-based cell survival [33]. In this study, we showed that inorganic sulfur treatment significantly reduces the expression of the mRNA, protein and active forms of ErbB2 and ErbB3. Although, the protein expression of pAkt is not affected by treatment of inorganic sulfur, the protein and mRNA expression of Akt were similarly reduced. Therefore inorganic sulfur seems to play a role in inhibiting the proliferation of MDA-MD-231 cells by inhibiting the EGFR-Akt pathway partly.
Various apoptosis-related genes, such as Bcl2, Bax, p53, and Fas, are involved in the apoptosis of tumor cells [34,35]. Bcl2 is an important gene that inhibits cell apoptosis, and Bax is an apoptotic protein [23]. Bax is generally found in the cytoplasm but translocates into the mitochondria by apoptotic stimuli causing apoptosis by inducing mitochondrial dysfunction [36]. The stimulation of the Akt pathway stimulates cell survival by inhibiting the translocation of Bax into the mitochondria [37]. Our results indicate that inorganic sulfur treatment induces cell apoptosis by decreasing the Bcl2/Bax ratio by increasing the expression of Bax without affecting Bcl2. Because treatment with inorganic sulfur was shown to reduce the mRNA and protein expresssion of inactive Akt without affecting the levels of the active form of Akt, inorganic sulfur apparently affects the apoptosis of breast cancer cells at least partly through the EGFR-Akt-Bax pathway.
The aforementioned results show that inorganic sulfur inhibits cell proliferation by inhibiting the expression and activation of EGFR and by increasing the expression of Bax in MDA-MD-231 human breast cancer cells. More research is needed to investigate the independent anti-carcinogenic effects of inorganic sulfur, which differ from those of organic sulfur.



 XML Download
XML Download