

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Anthocyanidins are bioactive flavonoid phytochemicals that are commonly found in edible plants. However, the stability of these compounds is easily affected by structural modifications with hydroxyl, methoxyl, carbonyl and acyl groups. Most anthocyanidins are turned into anthocyanins by glycosylation. The colors of purple sweet potato (PSP) are primarily achieved by the presence and form of anthocyanins. Due to the lack of toxicity, unique color, and nutritional and health benefits, the PSP has been regarded as a valuable source of food coloring and a healthy food.
Many studies have demonstrated that PSP anthocyanin (PSPA) has multiple physiological properties. In an in vitro active oxygen model, PSPA was found to possess strong free-radical scavenging activity that was concentration-dependent [1]. PSPA can inhibit the mutation of Salmonella typhimyrium TA98 [2]. Additionally, it was reported that PSP extracts can protect the liver from hepatotoxicity induced by paracetamol [3]. It was also found that PSPA offers protection against D-gal-induced structural damage and inflammatory responses [4], and can inhibit the overgrowth of bacteria such as Staphylococcus aureus and Escherichia coli [5]. PSPA extracts were found to have anti-tumor functions. Treatment with 5% PSPA may reduce rectal cancer development in mice [6], induce the apoptosis of cultured human stomach cancer cells and prevent the growth of stomach cancer in mice [7]. Moreover, diacylated anthocyanin derived from Ipomoea (I.) batatas cv. Ayamurasaki was found to have anti-hyperglycemic effects [8]. Significant protection against the development of spontaneous hypertension has also been observed in mice treated with plant-derived anthocyanin-rich compounds such as PSPA [9]. Despite these encouraging data, the studies mentioned above used different extraction method, so the color and activities of the extracts differ both in vitro and in vivo.
In a previous paper, we prepared PSPA extracts using a newly developed technique called microwave baking (MB) with acidified electrolyzed water (AEW). The resulting extracts had strong 2, 2-diphenyl-1-picrylhydrazyl (DPPH) free radical scavenging and anti-lipid peroxidation activities [10]. Based on our original study, the crude extracts were further purified in the present investigation to enable evaluation of their antioxidant, hypoglycemic, and anti-neoplastic activities in vivo.
Materials and methods
Materials
PSP (I. batatas) cv. Ayamurasaki was purchased from the Xuzhou Sweet Potato Research Institute, Chinese Academy of Science (Xuzhou, Jiangsu Province, China). Macroporous AB-8 resin was purchased from Shanghai Lan-Ji Science Technology Development Co. Ltd. (China). All kits used for this study were purchased from Nanjing Jian-Cheng Bioengineering Institute (China). Streptozocin (STZ) was purchased from Sigma Chemical Co. (USA).
Fifty 3-wk-old male Kunming ablactation mice [18-22 g body weight (BW)] and 50 Specific Pathogen Free (SPF)-grade ICR mice (25 males and 25 females, 18-22 g BW) were purchased from the Experimental Animal Center of Soochow University (Suzhou, China). This study was approved by the Institute Review Board of Soochow University (IRB #12-02-0040).
Sample of PSPA preparation and purification
PSPA was extracted as previously described [10] with some modification. PSPs were heated in a microwave oven for 15 min. The cooked potatoes (1 kg approximately) were crushed and suspended in acidic electrolyzed water (2 L, pH = 3.0). The suspension was cooled to 0℃ for 12 h without stirring. To remove the solid residue, the suspension was centrifuged at 12,000 rpm and 4℃ for 20 min on Avanti J-30I Centrifuge (Beckman, USA). Then the extracts were concentrated to remove water using a RE-5220 rotary evaporator (YaRong Biochemistry Instrument Factory, Shanghai, China). Through the Spray Dryer (Shanghai Trustech Co. Ltd., China) can transform into powder samples. The crude sample was dissolved in distilled water (1:5, W/V), and the solution loaded onto a solid phase extraction cartridge (AB-8 resin). The purified water was firstly eluted and the aqueous phase was discarded, 70% ethanolic water was secondly eluted and the ethanol-H2O eluent was collected. The eluent was concentrated to remove ethanol using a RE-5220 rotary evaporator (YaRong Biochemistry Instrument Factory, Shanghai, China). The concentrated aqueous elute was lyophilized on vacuum freeze dryer (Thermo, USA) for further evaluation.
PSPs were heated in a microwave oven for 15 min. The cooked potatoes (1 kg approximately) were crushed and suspended in acidic electrolyzed water (2 L, pH = 3.0). The suspension was cooled to 0℃ for 12 h without stirring. To remove the solid residue, the suspension was centrifuged at 12,000 rpm and 4℃ for 20 min prior to application onto an Amberlite AB-8 column. The column was rinsed with water and the extracts were eluted with a solution of water/ethanol (7:3 v/v). The eluent was concentrated in vacuo, freeze-dried and referred to as the crude PSPA extracts.
Determination of PSPA color value
The PSPA color was quantified using the spectrophotometric method previously described by Francis [11]. When the concentration of PSPA was 0.011% in aqueous solution, the ultraviolet (UV) spectrum was scanned from 200 to 600 nm in a 1-cm-thick absorption cell on U3000 Spectrophotometer (Hitachi Co. Ltd., Japan). Absorbance at the maximum absorption wavelength was measured. Color values for the PSPA extracts were calculated using the following equation:
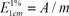
in which 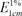 is the color value for the 1% extract in the 1-cm colorimetric cell at the maximum absorption wavelength, A is the absorbance, and m is the amount (0.2-0.7 g) of sample analyzed.
is the color value for the 1% extract in the 1-cm colorimetric cell at the maximum absorption wavelength, A is the absorbance, and m is the amount (0.2-0.7 g) of sample analyzed.
is the color value for the 1% extract in the 1-cm colorimetric cell at the maximum absorption wavelength, A is the absorbance, and m is the amount (0.2-0.7 g) of sample analyzed.Assay of total flavonoid (TF) contents
Weighting 10.0 mg constant weight rutin standard (Beijing Aoke Biotechnology Co. Ltd., China), dissolving it use 98% ethanol, then constanting volume to 50 mL, so we have 200 µg/mL rutin standard solution. Accurately draw rutin standard solution 0, 10, 20, 30, 40, 50 µL in the absorption cell, and then followed by adding 70% ethanol solution 50, 40, 30, 20, 10, 0 µL. Numbered the absorption cells 0, 1, 2, 3, 4, 5. To each standard, a 10% sodium nitrite solution (National Medicine Group Chemical Reagent Co. Ltd., China) (10 µL) was added. The solutions were shaken and left standing for 6 min. Next, 10 µL of a 20% aluminum nitrate solution (National Medicine Group Chemical Reagent Co. Ltd., China) were added before the mixture was shaken, and 50 µL of a 2 mol/L NaOH solution were added. Finally, 200 µL of a 60% ethanol solution was added to each standard. The mixtures were shaken and left standing for 10-15 min. Using the blank ("0") as the reference, absorbance was measured at 510 nm to generate the rutin standard solution curve. Least squares regression was subsequently used to evaluate the relationship between the absorbance values and rutin standard concentration.
An UV spectrophotometric method was used to determine TF contents expressed as rutin equivalents [12]. The samples (10.0 mg each) were dissolved in 10 mL of 60% ethanol, and 1 mL of the solution was added to a test tube. Absorbance was measured at a wavelength of 510 nm on U3000 Spectrophotometer (Hitachi Co. Ltd., Japan), and TF concentrations were determined using the standard curve. The analyses were done in triplicate.
Evaluation of total phenol (TP) contents
Gallic acid (National medicine group chemical reagent Co. Ltd.,China) (10.0 mg) was dried at 120℃ for 1h and dissolved in 100 mL water, to produce a 100 µg/mL gallic acid standard solution. Aliquots of the gallic acid standard solution (0.0, 0.1, 0.2, 0.4, 0.6, 0.8, and 1.0 mL) were transferred to 10 mL graduated test tubes (Sangon Biotechnological Co. Ltd., Shanghai, China). Next, 5.0 mL of distilled water and 0.5 mL Folin-Ciocalteu phenol reagent (National Medicine Group Chemical Reagent Co. Ltd., China) were added. The solution was shaken for 1 min before 1.5 mL of a 20% Na2CO3 solution was added. The mixture was then incubated for 2 h at room temperature. Using the blank standard, absorbance was measured at 760 nm to generate the standard curve.
An UV spectrophotometric method was used to determine the TP contents as gallic acid equivalents [13]. The samples (10.0 mg each) were dissolved in 10 mL of distilled water and 1 mL of the solution was transferred to a test tube. Absorbance was measured at wavelength of 760 nm on U3000 Spectrophotometer and TP concentration was calculated according to the standard curve.
Evaluation of antioxidant activity in aged mice
Twelve-mo-old male Kunming mice (50.36 ± 3.76 g BW) were obtained. The animals were randomly divided into five groups: the control group treated with 0.4 mL physiological saline, a positive group treated with 100 mg/kg BW of vitamin C or E (National Medicine Group Chemical Reagent Co. Ltd., China); three PSPA groups treated with low, medium, or high doses (100, 500 and 1,000 mg/kg BW, respectively) of PSPA; and a 5-month-old control group treated with 0.4 mL physiological saline. All the groups were treated for 30 d, and blood was collected from the eyeballs on 31th day. The serum was obtained at 4000 rpm for 20 min on the centrifuge (Beckman Coulter, Inc., USA). The total antioxidation capacity (T-AOC) as well as the levels of superoxide dismutase (SOD), malondialdehyde (MDA), and glutathione peroxidase (GSH-PX) in the serum were measured using the appropriate commercial kits. The assays were performed in triplicate. The data are expressed as the mean values and standard deviation (SD).
Assay to monitor hyperglycemia attenuation
Fifty 3-wk-old male Kunming ablactation mice (18-22 g BW) were maintained under controlled conditions (18-25℃, 50-80% humidity, and a 12-h light/dark cycle) with free access to rodent food (Suzhou Experimental Animal Food Co. Ltd., Suzhou, China) and tap water. The mice were divided into five groups. The control group (I) was treated with rodent food. The PSPA ig group (II) was treated with normal forage and 1.0 g PSPA/kg BW in 0.5 mL water was poured into the stomach daily by using special syringe (Sangon Biotechnological Co. Ltd., Shanghai, China). The PSPA po group (III) was treated with normal forage mixed with PSP extract (10 g/kg) as previously described [7]. Group IV was treated with a high-saturated fat, high-sugar diet (18% lard, 20% sugar, 3% egg yolk, and 59% basal) and PSPA. Finally, group V was treated with a high-saturated fat, high-sugar diet. All mice were fed for 4 wk according to their treatment group and BW was recorded weekly. After 4 wk, each mouse was weighed, injected with STZ (100 mg/kg BW), and then returned to their feeding regimen for another week. At the end of the feeding period, all the animals were fasted for 12 h. Blood samples were then collected by amputating the tail, blood glucose concentrations were measured using commercially available kits (BIOSINO Biotechnology and Science INC, China), and BW was recorded. Finally, all mice were anesthetized with pentobarbital sodium (National Medicine Group Chemical Reagent Co. Ltd., China) (35 mg/kg) and euthanized by cervical decapitation, and their hearts, livers, kidneys, and pancreas were excised and weighed. The organ index was calculated using the following formula:
Pancreas and islet tissues were fixed at 10% buffered formalin overnight and then immersed in 70% ethanol for storage at room temperature. The tissues were embedded in paraffin. Sections were cut at 5µm on positively charged slides. The section were stained with hematoxylin and eosin (National Medicine Group Chemical Reagent Co. Ltd., China) (HE) using conventional methods.
The food intake and water intake are obtained through the offered amount minus the surplus amount every day.
Evaluation of tumor development inhibition
Fifty SPF-grade ICR mice (25 males and 25 females, 18-22 g BW) were acquired from the Experimental Animal Center of Soochow University. The mice that were implanted with mice S180 anal sarcoma cells were provided by Nanjing Kai-Ji Biotechnology Development Co. Ltd (Nanjing, China). All animals were randomly assigned to five groups (n = 10 per group): a blank control (treated with 0.4 mL normal saline), negative control (implanted with tumor cells), and three treatment groups (implanted with tumor cells) given 100, 500, or 1,000 mg/kg BW PSPA.
During the experimental period, all animals had free access to five kinds of food (Table 4) and tap water. BW was recorded daily along with the numbers and sizes of sarcomas observed. The day after two successive weeks of treatments, the BW of each group was measured and blood was collected by removing the eyeballs. Entire sarcomas were removed from each mouse to measure tumor inhibition rates calculated with the following formula:
in which C is the average tumor weight for the negative control group (g) and T is the average tumor weight for the treatment group (g).
The rate of BW changes was evaluated with the following formula:
in which W is BW 2 wk after inoculation with the tumor cells (g) and W0 is the BW before inoculation with the tumor cells.
SOD, MDA, and GSH-PX levels were measured with the appropriate commercial kits.
Results
PSPA color values and composition after purification
PSPA extracts had strong absorption between 269 and 360 nm (Fig. 1) due to the characteristics of the acyl-anthocyanin structure. In the visible region (528 nm), the highest absorbance value was 0.335. The calculated color value of the sample was 30.15.
In the present study, rutin and gallic acid were used as reference standards to determine TF and TP contents, respectively, in the extracts. Two standard curves are quantities of rutin and gallic acid as abscissae and their absorbance values at 510 and 760 nm as ordinates, respectively, with the following regression equations: y = 0.01297x + 0.00429 and y = 0.01389x + 0.03654. The rutin or gallic acid concentrations and the absorbance values had a positive linear relationship. Using the linear regression equations, TF and TP concentrations were calculated as 932.5 mg/g and 140.789 mg/g, respectively. Based on these results, it appeared that anthocyanin extracted from PSP contained high TF and TP levels.
Effect on antioxidants on aged mice
Effect on the serum T-AOC contents
The PSPA-treated group had significantly higher T-AOC levels than the saline-treated control group. 100 mg/kg BW PSPA group was similar to the Vitamin C (VC) group in the effect on the T-AOC, while 1,000 mg/kg BW PSPA group was two-fold of the control (Table 1), showing that PSPA can enhance T-AOC in the serum of aged mice. The enhancement of T-AOC by treatment with PSPA showed dose-dependent.
Anti-aging effects
In the aged mice, PSP extracts significantly decreased serum MDA levels, and increased SOD and GSH-Px activities. The effect of treatment with low PSPA concentrations was similar to that elicited by the same amount of Vitamin E (VE). When the PSPA concentration was 500 mg/kg, the total SOD and GSH-Px activities increased by 23.94% and 28.34%, respectively, whereas the MDA level decreased by 20.64%. When the PSPA concentration was 1,000 mg/kg, the enzyme activities and inhibition of MDA production in the 12-mo-old mice was equal to that of the 5-mo-old animals, indicating significant retardation of the aging process (Table 2). These results suggest that PSPA confers protection against aging by increasing antioxidant activities.
Effects on STZ-treated diabetic mice
Plasma glucose
PSPA prevented the development of hyperglycemia to a significant extent in STZ-treated diabetic mice. Blood glucose levels of the two control groups (I and V) were clearly higher than those of the other three groups (II, III, and IV). Furthermore, the blood glucose concentrations increased over time. Values for the group fed a normal diet increased from 11.26 ± 3.338 to 18.57 ± 8.069 mmol/L while those for the high-saturated fat, high-sugar diet group increased from 13.71 ± 7.637 to 20.0 ± 9.518 mmol/L. In contrast, values for the animals fed the high-saturated fat, high-sugar diet and PSP did not change significantly. Glucose values for the treatment groups II, III decreased with time (Table 3). These results suggested that animals fed the diets containing high levels of saturated fat and sugar easily succumbed to hyperglycemia. The PSP extract significantly reduced plasma glucose levels in STZ-treated mice, but further studies are necessary to elucidate the underlying mechanisms of action.
Pancreatic morphology
Microscopic appearance of the pancreatic cells was normal. The pancreatic island was elliptical or circular while the boundary was clear and no membrane envelope was present. No hemorrhagic necrosis, interstitum vascular lesions or fibrosis, cell degeneration, or lymphocytic infiltration were observed.
Effect on BW and organ index
Control mice and ones fed a high-saturated fat, high-sugar diet lost significant amounts of weight 1 wk after STZ injection. When the animals were treated with PSP either via incorporation into the diet or intragastric delivery, no significant weight loss was observed (Table 3). These results indirectly indicate that PSPA can significantly inhibit STZ-induced diabetes and impair BW loss in mice. The mean organ indices for the liver and kidneys of the treatment groups were significantly lower than those for the control and high-saturated fat, high-sugar diet groups. However, no significant difference in the organ indices for the heart and spleen were observed between the groups (Table 4).
The present study showed that PSPA can significantly inhibit increases in plasma glucose levels in a dose-dependent manner. Over time, the hypoglycemic effect was even more significant. Diabetic mice showed growth retardation due to their high plasma glucose concentrations, but treatment with PSPA helped reverse this condition. However, PSP extract did not greatly affect major organ indices, possibly indicating that the extracts did not exert protective effects on those organs. The increased kidney organ index for the control and high-saturated fat, high-sugar diet mice may have resulted from renal injury.
Effect on tumor growth inhibition
Inhibition of implanted S180 cell growth by PSP extract in mice
Tumors were observed 5 d after implantation of S180 cells in the negative control group, and 1 wk after implantation in the treatment group (mortality rate was < 5% for both groups). The mean BW of mice in the negative group was lower than that of the PSPA group a week after S180 cell implantation (Table 5). The low, moderate, and high doses of PSP extract all inhibited tumor growth in the mice. When the PSP concentration reached 500 and 1,000 mg/kg BW, the inhibition rates were 43.28 and 68.03%, respectively.
Effect on SOD, GSH-PX, and MDA serum activities
In the PSPA groups, SOD and GSH-PX activities were somewhat lower while the MDA level was slightly increased compared to the blank control group, indicating that lipid peroxidation increased slightly in the experimental group. In contrast, the SOD and GSH-PX activities were higher while the MDA level were lower in the experimental group compared to the negative control group, indicating that PSPA prevented lipid peroxidation (Table 6). These findings suggest that PSPA could inhibit sarcoma growth in mice with the highest inhibition ratio of 68.03%. Due to increased SOD and GSH-PX activity as well as decreased MDA levels, the inhibitory effect of PSPA may be achieved by enhancing antioxidant activity in the mice.
Discussion
The intense purple color of the PSP flesh is thought to be due to anthocyanin accumulation. Anthocyanin extracted from PSP as a food colorant has numerous biological functions such as antioxidant, anti-aging, anti-tumor, and antimutagenic capacities. However, different PSP cultivars have remarkably different anthocyanin contents and biological activities. Thus, finding an economical and convenient way to acquire food colorant derived from PSP has become increasingly important.
In a previous study, compositions of four different PSP cultivars were determined by high-speed counter current chromatography with crude materials extracted in a solution of water and hydrochloric acid [14]. In the present investigation, we prepared samples by MB and AEW (MB-AEW), and the suspension was applied to an Amberlite AB-8 cartridge for further purification. This new MB-AEW technique not only prevents the degradation of non-acylated anthocyanins, but does not introduce new ions such as acetates into the extract. The color value of PSP extract was 30.15 after purification. The TF content was 93.25% (expressed as rutin equivalents) and the TP content was 14.08% (expressed as gallic acid equivalents).
Several studies have been performed to evaluate the in vitro antioxidant activity of PSP. The most common experimental method to measure antioxidant activity has been to assess DPPH radical scavenging activity [15,16] along with ORAS and ABTS radical scavenging activities [17]. Many biological functions of PSP such as memory enhancement [16], neuroprotection, and anti-apoptotic effects [18] may be associated with antioxidant properties of the plant. Assessment of antioxidants in vivo indicated that PSPA could significantly retard the aging process in older mice. Administration of PSP extracts to 12-mo-old mice for 1 mo resulted in an in vivo anti-aging index equal to that of 5-mo-old mice. Matsui et al. [8] described the antihyperglycemic effect of diacylated anthocyanin derived from I. batatas cv. Ayamurasaki. When diacylated anthocyanin (100 mg/kg) was administered after treatment with maltose (2 g/kg), the maximal blood glucose level observed at 30 min was significantly decreased by 16.5% [19]. In our study, no significant differences BW, food intake, water intake, or pancreatic morphology were observed between the control and PSPA extract-treated groups. PSPA also notably lowered plasma glucose in STZ-treated mice.
Acylated anthocyanins in PSP were previously found to be easily absorbed in rats [20], and could be used to promote good health due to this characteristic. Flavonoid from Cirsium japonicum DC has been found to inhibit tumor activity and regulate the immune response in mice with S180 and H22 tumors [19]. Hagiwara et al. [6] confirmed that administration of a 5.0% PSP pigment solution can prevent colorectal cancer in mice. Additionally, Hayashi et al. [7] confirmed that PSP anthocyanins can induce programmed cell death in cultured human gastric cancer cells in mice with a growth inhibition rate of 47.6%. In our study, PSPA effectively suppressed tumor growth with the highest inhibitory rate of 68.03%. Furthermore, PSPA consumption elevated GSH-PX and SOD levels and lowered MDA levels, and may indicate that inhibition of tumor growth was achieved through enhancement of antioxidant activities. Although this purified extract may have many health benefits, it is a mixture that includes many different types of anthocyanins. Further studies are necessary to isolate and prepare individual components to determine which one possesses the beneficial biological properties observed in our study.







 XML Download
XML Download