

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
The color of skin and hair results from the synthesis of melanin in melanocytes [1]. Melanin is generated by ultraviolet (UV) light and acts as a direct defense against UV radiation [2]. However, excessive melanin synthesis causes hyperpigmentation disorders, including melasma, freckles, lentigo, and other hyperpigmentation syndromes [3]. Melanosomes are specialized lysosome-related organelles that synthesize and store melanin pigments. Mature melanosomes are transferred from melanocytes to keratinocytes, leading to movement to the skin surface [4]. Several well-known enzymes, such as tyrosinase, tyrosinase-related protein (TRP) 1, and TRP2, play vital roles in melanosome biogenesis, and these proteins catalyze melanin synthesis [45]. Mutations in tyrosinase and TRP1 are involved in human pigment disorders, such as oculocutaneous albinism (OCA) 1 and OCA3, respectively [6].
Microphthalmia-associated transcription factor (MITF) is the core transcription factor involved in the expression and transport of melanosome component proteins [7]. The pivotal role of MITF is to control expression of melanogenic enzyme proteins including tyrosinase, TRP1, and TRP2, in addition to its role in the transport of melanosomes to the dendritic tips [38]. α-Melanocyte stimulating hormone (α-MSH) binds to melanocortin-1 receptor 1 (MC1R) and sequentially stimulates MITF expression [9]. MC1R triggers the cAMP pathway when activated, leading to cAMP-dependent transcriptional activation of MITF expression [10]. Although the role of glycogen synthase kinase 3 beta (GSK3β) in the induction of melanogenesis remains controversial, recent studies have reported that the inhibition of GSK3β phosphorylation induces the degradation of β-catenin and inhibits the transcriptional and protein expression of MITF and melanogenic enzyme proteins [11121314]. Melanogenesis stimulated by α-MSH increases the phosphorylation of GSK3β, leading to phosphorylation at Ser675 and stabilization of β-catenin protein. Subsequently, β-catenin accumulates in the cytoplasm by escaping ubiquitination-dependent proteasomal degradation, but relocalizes to the nucleus where it works in concert with cAMP response element binding protein (CREB) on the MITF promoter [15]. Therefore, exposure to GSK3β-specific inhibitors results in the induction of melanin synthesis and expression of tyrosinase and MITF [13].
Various skin depigmentation agents, such as arbutin, kojic acid, and hydroquinone, reduce melanin production [16]. However, these agents induce toxic, potentially carcinogenic side effects, and skin irritation [17181920]. Thus, whitening agents from natural sources are preferred and tend to dominate the cosmetic market [21]. Sageretia thea, belonging to the Rhamnaceae family, has long been recognized as a medicinal herb in Korea and China [22232425]. Several studies have demonstrated that the leaves of Sageretia thea have antioxidant activity [2223] and have been used to treat itching, boils, and inflammation of the skin caused by lacquer poison in China [25]. Although some studies have demonstrated the biological functions of Sageretia thea leaves, few studies have investigated the chemical and biological composition of Sageretia thea fruits. Notably, the fruit constitutes an important source of active metabolites against skin aging; thus, several studies have reported on melanogenic-inhibitory activities and fruit constituents [262728]
In the present study, we examined the anti-melanogenesis effect of Sageretia thea fruit and its underlying molecular mechanisms in B16F10 cells in an effort to develop new, safe, and effective skin lighteners from natural sources. Various solvent fractions from the crude extract were examined to obtain fractions with greater anti-melanogenic effects, and a gas chromatograpy-mass spectroscopy (GC-MS) analysis was conducted to identify the potential ingredients from the active solvent fractions. We identified the major anti-melanogenic compounds (methyl linoleate and methyl linolenate) from the n-hexane fraction, and we measured melanin content and intracellular tyrosinase inhibitory activity in B16F10 cells. Furthermore, we determined the levels of tyrosinase, TRP1, MITF, Akt/p-Akt, GSK3β/p-GSK3β, and β-catenin by western blotting and elucidated the biological mechanisms underlying the inhibitory effect of melanogenesis in the n-hexane fraction from Sageretia thea fruit (HFSF), as well as those of methyl linoleate and methyl linolenate.
MATERIALS AND METHODS
Reagents and antibodies
Methyl linoleate, methyl linolenate, α-MSH, L-DOPA, sodium hydroxide (NaOH), mushroom tyrosinase, arbutin, kojic acid, resveratrol, and 6-bromoindirubin-3'-oxime (BIO) were obtained from Sigma-Aldrich (St. Louis, MO, USA). Antibodies recognizing Akt, p-Akt, GSK3β, p-GSK3β, and β-actin were obtained from Cell Signaling Technology (Beverly, MA, USA). Anti-β-catenin antibody was purchased from BD (Franklin Lakes, NJ, USA), and anti-MITF, anti-tyrosinase, and anti-TRP1 antibodies were obtained from Santa Cruz Biotechnology (Santa Cruz, CA, USA).
Plant material extraction and fractionation
Sageretia thea fruit was harvested from Seogwang, Jeju Island, South Korea in May 2014. Botanical samples were identified taxonomically by Dr. Hui Kim, Mokpo National University, Korea. Sageretia thea fruit (10 kg) was extracted with 80% aqueous methanol for 24 h. After concentrating the methanol extract, the crude extract (1.6 kg) was partitioned with n-hexane, chloroform, ethyl acetate, n-butanol, and water. The fractions were generated using 18 g of n-hexane, 3.6 g of chloroform, 20.9 g of ethyl acetate, 135.0 g of n-butanol, and 1,422.5 g of water. The n-hexane fraction was further partitioned using a silica gel column and Sephadex LH-20 column. In total, 18 fractions (H-1-H-18) were obtained and monitored by thin-layer chromatography. After fraction H-3 (1.47 g) was partitioned using a silica gel column, nine fractions (H-3-1-H-3-9) were obtained and monitored by TLC.
GC-MS analysis
GC-MS analysis was performed with a Shimadzu model QP-2010 (Kyoto, Japan) in EI mode (70 eV) using a capillary Rtx-5MS column (30 m × 0.25 mm, 0.25 µm film thickness). A 1 µL volume was injected with a 1:10 split ratio at temperatures of 250℃ at the inlet and 290℃ at the interface. An initial temperature of 60℃ for 2 min was increased to 250℃ at a rate of 5℃/min and then increased to 310℃ at a rate of at 8℃/min for 12 min. Mass range was scanned from 40 to 500 amu. Data were identified using the WILEY9 and NIST05 libraries.
Cell culture and cell viability
B16F10 mouse melanoma and human dermal fibroblast cells were kindly provided by Dr. Nam Ho Lee and Dr. Moonjae Cho, Jeju National University, Korea, respectively. The cells were cultured in DMEM with 10% heat-inactivated fetal bovine serum (FBS) and 1% penicillin-streptomycin (Invitrogen, Grand Island, NY, USA) in a 5% CO2 atmosphere at 37℃. Cell viability was examined using the MTT assay (Amresco Inc., Solon, OH, USA). Briefly, cells were treated with various concentrations of samples. After 48 h, the MTT solution was added to the cells for 4 h. After solubilizing the formazan crystals with DMSO, the absorbance at 570 nm was detected using a microplate reader (Tecan, Salzburg, Austria).
Melanin content
Using a slightly modified version of the method described previously, we conducted an assay to determine the melanin content [29]. Cells were treated with various concentrations of HFSF in the absence or presence of α-MSH for 48 h. After harvesting the cells by centrifugation, the pellets were dissolved with 1 N NaOH and 10% DMSO at 80℃ for 30 min, and the absorbance was measured at 475 nm using the microplate reader.
Tyrosinase activity
Intracellular tyrosinase activity assays were performed according to a slightly modified version of the method described previously [16]. Cells were treated with various concentrations of HFSF in the absence or presence of α-MSH for 48 h. The cells were homogenized with cold RIPA buffer and protease inhibitor cocktail. The protein content was normalized with a BSA protein assay kit (Pierce, Rockford, IL, USA) and then incubated with L-DOPA (2 mM) at 37℃ for 30 min. Direct tyrosinase activity was determined using a mushroom tyrosinase solution. Samples of varying dilutions were incubated with L-DOPA (2 mM) at 37℃ for 20 min. Tyrosinase activity was detected using a microplate reader at an absorbance of 490 nm. in situ tyrosinase activity was assessed as described previously [30]. Cells were treated with various samples in the absence or presence of α-MSH for 48 h. The cells were fixed with 4% paraformaldehyde for 40 min. After permeabilizing the cell with 0.1% Triton X-100, cells were incubated in L-DOPA (2 mM) for 2 h at 37℃. The cells were examined using a microscope (Olympus, Essex, UK).
Western blotting
Cells were treated with various concentrations of HFSF in the absence or presence of α-MSH for 48 h. The cells were homogenized with cold RIPA buffer with protease inhibitor cocktail and then normalized using a BSA protein assay kit. The cell lysates were separated by 12% SDS-PAGE and then transferred onto a polyvinylidene difluoride membrane. The membrane was blocked with 5% nonfat dried milk. After incubating the membrane with primary antibodies, the membrane was probed using horseradish peroxidase-conjugated goat anti-mouse or anti-rabbit IgG secondary antibodies for 30 min. Specific proteins were detected using the BS ECL Plus kit (Biosesang, Gyeonggi-do, Korea).
Statistical analysis
All data analysis was performed by one-way analysis of variance (ANOVA) using SPSS (SPSS Inc., Chicago, IL, USA). The data were analyzed as means ± standard deviation (SD). At least three independent experiments were conducted for each experiment. P values < 0.05 or < 0.01 were considered to indicate statistical significance.
RESULTS
The hexane fraction of Sageretia thea fruit inhibits α-MSH-induced melanogenesis
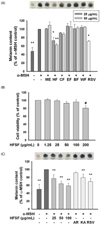
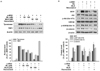
The Sageretia thea fruit crude 80% methanol extract and its n-hexane, chloroform, ethyl acetate, n-butanol, and water fractions at concentrations of 25 and 50 µg/mL were compared for their inhibitory activities against melanin production in α-MSH-induced B16F10 cells. The n-hexane fraction from Sageretia thea fruit (HFSF) with the highest inhibitory activity against melanin production was chosen among these fractions (Fig. 1A). As shown in Fig. 1B, no obvious toxicity in B16F10 melanoma cells was observed when the HFSF concentration reached 100 µg/mL. The HFSF affected growth of human dermal primary fibroblast cells when its concentration was ≥ 100 µg/mL (Fig. S1A). Thus, ≤ 100 µg/mL of the HFSF was used in subsequent experiments. This non-toxic HFSF treatment significantly suppressed melanin production in a concentration-dependent manner (Fig. 1C). Moreover, the anti-melanogenic effec of HFSF was superior to those of three common whitening agents named arbutin (2 mM), kojic acid (400 µM), and resveratrol (20 µM).
The HFSF suppresses intracellular tyrosinase activity and its expression
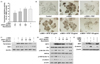
Tyrosinase induces the oxidation of tyrosine as a first step of melanogenesis [31]. Therefore, we treated B16F10 cells with the HFSF to examine the tyrosinase activity. α-MSH-stimulated tyrosinase activity was reduced in a concentration-dependent manner by the HFSF (Fig. 2A). In addition, when B16F10 cells were incubated with L-DOPA to detect in situ tyrosinase activity, we observed that HFSF reduced the staining in cells in the presence of L-DOPA (Fig. 2B). In contrast, no difference in mushroom tyrosinase activity was observed between treatment with or without the HFSF (Fig. S1B). These results indicate that cellular processes were modulated to decrease tyrosinase activity. Next, we examined the expression of melanogenic-related enzyme proteins by western blotting. As shown in Fig. 2C, treatment with the HFSF significantly diminished the levels of tyrosinase and TRP1 in a concentration-dependent manner. However, no change in TRP2 protein expression was detected (data not shown). These results indicated that suppression of melanogenesis with the HFSF was related to interrupting the upstream pathway that regulates the expression of tyrosinase and TRP1.
The HFSF regulates Akt/GSK3β/β-catenin signaling to inhibit MITF expression
We determined the effect of the HFSF on MITF and phosphorylation of GSK3β to elucidate the mechanism by which it inhibited melanin synthesis. The GSK3β/β-catenin signal pathway is closely related to melanin synthesis [32]. Inhibition of GSK3β and concurrent accumulation of β-catenin result in activation of the MITF promoter during α-MSH-stimulated melanogenesis and ultimately MITF target genes, such as tyrosinase, TRP1, and TRP2 [333435]. Previous reports suggested that activated Akt phosphorylates GSK3β and inhibits degradation of β-catenin [1433]. As shown in Fig. 2D, α-MSH treatment significantly increased the MITF level, whereas co-treatment with HFSF had a concentration-dependent inhibitory effect on the MITF level. The data show that the level of GSK3β phosphorylation decreased, while the total GSK3β protein expression level remained unchanged. As expected, the expressions of phosphorylated Akt and β-catenin were decreased significantly in B16F10 cells treated with the HFSF (Fig. 2D). To confirm whether the expression of the β-catenin protein is directly regulated by GSK3β in HFSF-treated cells, the cells were treated with the GSK3β inhibitor BIO. The expression levels of MITF and β-catenin increased in response to co-treatment with 2 µM BIO compared to those of 100 µg/mL HFSF alone (Fig. 2E). These results indicated that the HFSF suppresses the expression of MITF via the Akt/GSK3β/β-catenin signal pathway, thereby diminishing the expression of melanogenic enzymes and ultimately reducing melanin production.
Methyl linoleate and methyl linolenate are major active constituents of the HFSF
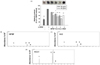
As the HFSF inhibited melanogenesis, we conducted bioassay-guided fractionation of the HFSF. The HFSF was further separated by silica gel column chromatography into 18 sub-fractions (H-1-H-18), using methanol as the mobile phase. The cells treated with sub-fraction H-3 inhibited melanin production the most among all of the sub-fractions (Table S1). H-3 was subjected to further fractionation into nine sub-fractions (H-3-1-H-3-9), which were evaluated for their inhibitory activity against melanin production. H-3-1 was the sub-fraction that inhibited melanin content most in α-MSH-stimulated B16F10 cells (Table S2). To compare the melanin content, H-3 and H-3-1 were tested at the same concentrations as the HFSF; the inhibitory activities of melanin production were enriched in sub-fraction H-3-1 (Fig. 3A). Moreover, these fractions inhibited in situ tyrosinase activity in a manner similar to that observed with the melanin content (Fig. S2). Next, we analyzed the chemical profiles of the HFSF, H-3, and H-3-1 to identify the constituents responsible for anti-melanogenesis. GC-MS analysis revealed that methyl linoleate (21.03 ± 6.73%) and methyl linolenate (20.97 ± 6.68%) were the major constituents of the HFSF, whereas methyl linoleate, methyl linolenate, and β-sitosterol were major constituents of H-3 with contents of 21.99 ± 0.97%, 23.77 ± 1.61, and 20.57 ± 2.24%, respectively (Table 1). Similarly, the major constituents of H-3-1 were methyl linoleate (39.24 ± 1.28%) and methyl linolenate (41.57 ± 5.25%), which were found in the HFSF, H-3, and H-3-1 and were enriched in sub-fraction H-3-1 (Fig. 3B–D). These results reveal that the anti-melanogenic activity of the HFSF was likely due to the presence of methyl linoleate and methyl linolenate.
Methyl linoleate and methyl linolenate downregulate MITF and tyrosinase-related proteins
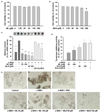
As methyl linoleate and methyl linolenate were major constituents of H-3-1, we examined the anti-melanogenic effect of methyl linoleate and methyl linolenate in α-MSH-stimulated B16F10 cells. Cell viability was not affected at concentrations ≤ 100 µM methyl linoleate or methyl linolenate. However, cell viability decreased to 73.70 ± 5.57% in response to 200 µM methyl linolenate, but not to 200 µM methyl linoleate (Fig. 4A and B). Furthermore, neither compound was toxic to human dermal primary fibroblast cells at 200 µM (Fig. S3A and B). Therefore, 100 µM, which showed no toxicity to either cell type, was chosen to determine the anti-melanogenic effects of methyl linoleate and methyl linolenate in B16F10 cells. As shown in Fig. 4C, α-MSH-stimulated melanin production was almost completely blocked by methyl linoleate and methyl linolenate. Similarly, methyl linoleate and methyl linolenate decreased α-MSH-stimulated intracellular tyrosinase activity and in situ tyrosinase activity in concentration-dependent manners (Fig. 4D and E). Moreover, the inhibitory activities of methyl linolenate on melanin production and intracellular tyrosinase activities were stronger than those of methyl linoleate. Notably, treatment with methyl linoleate and methyl linolenate showed no inhibitory effect on mushroom tyrosinase activity (Fig. S3C). As shown in Fig. 5A, methyl linoleate and methyl linolenate inhibited the expression of tyrosinase and TRP1 in α-MSH-treated B16F10 cells, suggesting that methyl linoleate and methyl linolenate inhibited α-MSH-induced intracellular tyrosinase activity by lowering expression of the melanogenic enzymes. Consistent with the HFSF treatment, methyl linoleate and methyl linolenate decreased MITF expression and Akt and GSK3 β phosphorylation, respectively (Fig. 5B). Moreover, decrease in the β-catenin level was detected in B16F10 cells treated with methyl linoleate and methyl linolenate. These results suggest that methyl linoleate and methyl linolenate suppressed melanin production by suppressing MITF in an Akt/GSK3β/β-catenin-dependent manner, resulting in decreased tyrosinase and TRP1 expression. Therefore, the anti-melanogenic effect of the HFSF was attributed to methyl linoleate and methyl linolenate. Taken together, our results suggest that suppressing MITF through the Akt/GSK3β/β-catenin pathway mediated the anti-melanogenic effect of the HFSF, which contains methyl linoleate and methyl linolenate as major constituents.
DISCUSSION
The fruits of Sageretia thea are purple and contain micronutrients [24]. Sageretia thea fruits contain a much higher level of anthocyanins than do blueberries [2436]. Anthocyanins are beneficial bioactive flavonoids known to reduce melanin production and mushroom tyrosinase inhibitory activity in α-MSH-stimulated B16F10 cells [37]. Therefore, we expected that Sageretia thea fruits would have a whitening effect derived from the presence of a large number of anthocyanins. However, we found little anthocyanin present in the n-hexane and ethyl acetate fractions of Sageretia thea fruits. Notably, in this study, bioassay-guided fractionation of the HFSF followed by GC-MS analysis identified methyl linoleate and methyl linolenate as major constituents. Interestingly, methyl linoleate and methyl linolenate isolated from Oxalis triangularis significantly block forskolin-induced melanogenesis by inhibiting cAMP production and tyrosinase promoter activity in B16 mouse melanoma cells [38]. However, no analysis of melanogenic protein expression levels was conducted in that study. In agreement with this previous report, we observed that treatment with methyl linoleate and methyl linolenate decreased melanin content. Furthermore, our data showed that methyl linoleate and methyl linolenate decreased the expression of MITF, tyrosinase, and the TRP1 protein and reduced intracellular tyrosinase activity in α-MSH-treated B16F10 cells, suggesting that the anti-melanogenic activities of methyl linoleate and methyl linolenate are attributed to inhibited transcriptional activation of MITF expression.
In the process of melanin synthesis, tyrosinase initiates melanogenesis via the oxidation of tyrosine and then yields L-DOPA and dopaquinone as the final common precursors [33]. Tyrosinase plays a role in regulating general melanin synthesis as a first and rate-limiting enzyme [31]. Additionally, melanin synthesis requires TRP1 and TRP2, which catalyze specific steps in melanogenesis [39]. Moreover, a strong intermolecular association between TRP1 and tyrosinase has been demonstrated to stabilize tyrosinase [40]. Our data indicated that the intracellular tyrosinase inhibitory activity of the HFSF was similar to that of resveratrol, a tyrosinase inhibitor positive control. When cells were treated with 100 µg/mL HFSF, the intracellular tyrosinase inhibitory activity of HFSF was similar to that of 20 µM of resveratrol, a positive control of tyrosinase inhibitor, as shown in Fig. 2A. Interestingly, the reduction of TRR1 expression with 100 µg/mL of HFSF was more pronounced than with 20 µM resveratrol, affecting stabilization and tyrosinase activity (Fig. 2C). Additionally, the HFSF contained γ-tocopherol (0.72 ± 0.15%) and ethyl linoleate (2.02 ± 0.19%), which have inhibitory effects on melanogenesis (Table 1) [3841]. Thus, the anti-melanogenic effect of the HFSF could be derived from the synergistic action of γ-tocopherol and ethyl linoleate, as well as methyl linoleate and methyl linolenate. Therefore, the HFSF itself could have potential as a whitening agent.
A relationship between Wnt/β-catenin signaling and MITF was demonstrated previously by physical interactions between the β-catenin protein and T-cell transcription factor/lymphoid enhancer binding factor, which directly binds and regulates the MITF gene to activate MITF target genes encoding tyrosinase, TRP1, and TRP2 [34]. Notably, α-MSH stimulates phosphorylation of GSK3β at Ser9 in a protein kinase A-dependent fashion resulting in inactivation of GSK3β. The attenuation of GSK3β activity facilitates stabilization of β-catenin [15]. α-MSH stimulation also stabilizes β-catenin by a dual mechanism in which p21-activated kinase 4 (PAK) induces the phosphorylation of β-catenin at Ser675 while blocking β-catenin phosphorylation at Ser33/37 [42]. The accumulated β-catenin in the cytoplasm is translocated into the nucleus where it combines with the MITF promoter contributing to transcription of MITF [15]. In addition, activation of Akt phosphorylates GSK3β at Ser9, which eventually inhibits degradation of β-catenin [1443]. Our data indicate that the HFSF attenuated phosphorylation of Akt and GSK3β, in other words, the inactive form of GSK3β decreased, leading to a reduction in β-catenin, and a decrease in MITF expression. Treatment with BIO, a kind of GSK3β inhibitor, restored HFSF-induced β-catenin inhibition. Similarly, we observed that inhibiting MITF with methyl linoleate and methyl linolenate was dependent on the Akt/GSK3/β-catenin pathway, and treatment with BIO reversed the reduction in β-catenin expression caused by methyl linoleate and methyl linolenate (data not shown). Therefore, the HFSF, methyl linoleate, and methyl linolenate inhibit β-catenin-mediated transcriptional activation of MITF through the Akt/GSK3β signaling pathway.
In conclusion, this study is the first to demonstrate that Sageretia thea fruit possesses anti-melanogenic activity without being toxic to B16F10 cells. The HFSF suppressed melanin content and intracellular tyrosinase activity by downregulating the expression of tyrosinase, TRP1, and MITF. Notably, the methyl linoleate and methyl linolenate were identified as major constituents of HFSF. We demonstrated that the underlying mechanism of anti-melanogenic activity of the HFSF, methyl linoleate, and methyl linolenate was the inhibition of MITF expression mediated by the Akt/GSK3β/β-catenin pathway. Hence, HFSF, methyl linoleate and methyl linolenate could be applied as whitening agents in cosmetics and in the clinic for treating hyperpigmentation disorders.

 XML Download
XML Download