

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
The early life stages of fish are potentially useful as an alternative experimental model [9] because embryonic stages are the most sensitive in the life cycle of the teleost [17,18]. In addition, fish have been used for animal welfare reasons. The main benefits of using zebrafish as a toxicological model over other vertebrate species are with regards to their small size, husbandry, and early morphology [13]. Furthermore, zebrafish embryos that are malformed can usually survive substantially past the time in which those organs start to function in healthy individuals [13]. For example, mutant zebrafish such as still heart, and slow mo [6], and toxicant-exposed embryos with heart abnormalities [2,14] survive well beyond 24 h when the heart normally begins to beat [16].
Because of the advantages of this in vivo system, we studied the developmental toxicities of some well-known environmental pollutants, cytochrome P4501A1 (CYP1A1) inducers, such as 3,3',4,4',5-pentachlorobiphenyl (PCB126), 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD), and 3-methylcholanthrene (3-MC), on zebrafish embryos. We also established an in vitro system using a human CYP1A1 promoter on a zebrafish liver cell line to detect the toxicity levels of aryl hydrocarbon receptor (AhR)-binding toxicants. The CYP1A1 inducers, AhR ligands, activated AhR [5]. Activated AhR, an α-class protein, dimerizes with a β-class protein, such as the AhR nuclear translocator, and the heterodimer can bind putative xenobiotic response elements (XREs, aka, AhREs, DREs) in the 5'-flanking region of the CYP1A1 [31]. The CYP1A1 promoter/enhancer was also previously isolated and characterized by Zeruth and Pollenz in zebrafish [32].
The main objective of this study was to demonstrate the potential for obtaining valuable and novel insights in chemical toxicology using the zebrafish as an alternative model vertebrate. We have also established a transient transgenic zebrafish liver cell line for AhR mechanism study.
Materials and Methods
Chemicals
PCB126 of 99.8% purity was obtained from Neosyn Laboratories (USA), TCDD of >99% purity was obtained from Supelco (USA), and 3-MC of 98% purity were obtained from Sigma-Aldrich (USA). PCB126, TCDD, and 3-MC were dissolved in 0.01% DMSO prior to use in order to obtain stock solutions 800 times the experimental concentrations. Each of the chemicals was administered to zebrafish larvae for 96 h to establish a concentration range for the toxicity and morphological changes.
Zebrafish maintenance
Fish were purchased from a local supplier. Adult zebrafish were raised and maintained on a 14 : 10 h light : dark cycle at 28.5℃ and were bred in tanks as described by Westerfield [30]. Mature fish were fed twice daily with a combination of Freshwater Aquarium Flakefood (TetraWerke, Germany) and live brine shrimp (San Francisco Bay Brand, USA). Care and treatment of the animals was conducted in accordance with guidelines established by the Institutional Animal Care and Use Committee, Seoul National University.
Median lethal concentration (LC50) and median combined adverse effect concentration (EC50)
To determine LC50 and EC50 levels and to determine the chemical exposure doses for the following experiments, we first performed acute toxicity tests using Blechinger's method [4]. As part of the acute toxicity tests, embryos were immediately exposed to DMSO (0.01%) or one of the chemicals (0-200 nM (67.2 mg/ml) for PCB; 0-155 nM (50 ng/ml) for TCDD; 0-50 µM (12.4 mg/ml) for 3-MC) for 96 h. Three replicate treatment groups (3 × 30 embryos) were exposed to each dose in 6-well polystyrene multi-well plates (10 embryos per well). Six or 12-well polystyrene multi-well plates (SPL, Korea) were silanized to minimize interaction of the solute with active sites on the walls as follows: Dimethyldichlorosilane (1 ml) in heptane (5%; Sigma-Aldrich, USA) was added to each well and exposed for 1 h rat room temperature. After the incubation period, the solution was removed and the plates were air-dried. Morphological observations were recorded, and the solutions were changed twice daily. Dead larvae were counted and removed. The average proportion of larvae corresponding to a given end point was calculated for each concentration.
Heart rate and hatching time
Five eggs were randomly distributed into each well of 12-well polystyrene multi-well plates, with 6 replicates (2.5 ml test solution per well). The multi-well plates were kept at 28.5℃, with a photoperiod of 14 : 10 h light : dark cycle. After 24 h, all dead embryos were removed and the number of living eggs was reduced to 20 to obtain an equal number of embryos prior to starting the subsequent experiments. The normal mortality rate during the first developmental stages was calculated to lie between 5% to 40% with OECD 212 [23]. Then five eggs were redistributed into each well (five eggs in four wells). Two concentrations for each chemical were used to determine LC50 and EC50 (0.4 and 100.0 nM for PCB126, 2 nM and 20 nM ng/mL for TCDD, 1 and 10 µM for 3-MC). At 48 h post fertilization (hpf), the heart is comprised of two chambers and beats regularly. The heart rate was calculated by direct observation of the heartbeat for 10 sec. At 48 hpf, the embryos are able to hatch. The number of hatched prolarvae was recorded every 2 h until 80 hpf. A prolarva is considered hatched when the entire body (from tail to head) is out of the chorion. The hatching rate was calculated for each multi-well plate as the percentage of hatched larvae per plate. Then the number of hatched embryos in each replicate was pooled to calculate the mean hatching time (HT50) by Fraysse's method [9].
Plasmid construction
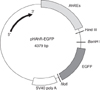
The human AhR-regulated reporter plasmid, phAhRE-EGFP, was described in detail in our previous study [26]. In brief, it was constructed by fusing a portion of the two consensus aryl hydrocarbon response element (AhRE) sequences, and this oligonucleotide was ligated into pEGFP vector using the Ase I and Hind III sites (Fig. 2).
Zebrafish liver cell culture
We cultivated adult zebrafish liver (ZFL) cells (ATCC CRL-2643) in ZFL medium consisting of Leibowitz L-15, Dulbecco's Modified Eagle's Medium, and Ham's F12 (50 : 35 : 15) supplemented with 10 mg/ml insulin, 5% fetal bovine serum, and 50 ng/ml epidermal growth factor at 28.0℃, as described by Ghosh et al. [10]. Cells were grown in 75 mm tissue culture plates and the culture medium was changed every other day.
Transient transfection
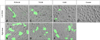
One day before transfection, 1.0 × 106 ZFL cells per well were plated (6-well polystyrene multi-well plates) in 500 µl growth medium without antibiotics, resulting in 90-95% confluence at the time of transfection. Lipofectamine 2000 (Invitrogen, USA) was used according to the manufacturer's recommendations for transient transfection studies. Cell survival rates after chemical exposure were calculated by the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) tetrazolium assay [20]. According to MTT assay results, the treatment concentrations of each chemical were determined (data not shown). Then cells were treated with two concentrations of each agent, namely 500 nM and 10 µM for PCB126, 20 nM and 200 nM for TCDD, 1 µM and 10 µM for 3-MC, for 24 h in 5 ml serum-free L-15 medium and then washed before conducting the bioassays. All experiments were carried out in triplicates. The cells were observed using a Nikon C1si spectral imaging confocal system (Nikon, Japan) with a laser that emits at 488 nm. The display merged the differential interference contrast image with the enhanced green fluorescent protein (EGFP) expression image.
Results
Embryotoxicity of PCB126, TCDD, and 3-MC
Prior to the experiments with transgenic zebrafish, PCB126, TCDD, and 3-MC were administered to naive zebrafish larvae for 96 h to establish a concentration range for the CYP1A1 expression studies and to determine the acute toxicity of PCB126, TCDD, and 3-MC in the early larval stage of zebrafish (Fig. 1). The predominant effects observed after PCB126 exposure were pericardial edema, swollen yolk sac, trunk abnormalities and contorted tail (Fig. 1A). As shown in Fig. 1C, vascular impairment was detected, such as irregular arrangement of the caudal artery and vein. The main toxic manifestations of the well-known AhR ligand, TCDD, reported in our acute toxicity tests included brain hemorrhage, somite irregularity, elongated and unlooped heart, pericardial edema, no swim bladder inflation, swollen yolk sac and lower jaw shortening (Figs. 1E-H). 3-MC predominantly caused elongated and unlooped heart, pericardial edema, no swim bladder inflation, swollen yolk sac, and lower jaw shortening (Figs. 1I and J). These data suggested that AhR ligands might cause gross morphological differences, and accurately predicted sites of chemical-induced toxicities, as summarized in Table 1.
The LC50 and EC50 and their 95% confidence intervals were calculated in Table 2 respectively. Apart from gross morphological differences, sub-lethal effects (heart rate and hatching time disturbance) were observed following exposure to PCB126, TCDD, and 3-MC. At 48 hpf, the heart consists of chambers and presents a regular heart rate. This sub-lethal end point was calculated by direct observation of the heartbeat for 10 sec. The heart rate is described by discrete values. The mean values of heart rates for PCB126, TCDD, and 3-MC respectively are reported in Table 2. The recorded values ranged from 19 to 24 beats/10s. The heart rate of each two concentrations of three model toxicants indicated no statistically significant differences (p < 0.05) at 48 hpf zebrafish. For all groups, 100% of the embryos hatched at 72 hpf and there was no time lag between the first and the last hatching. Only high concentration 3-MC exposure delayed hatching, as measured by HT50 estimation, compared to low concentration and the negative control.
Exposure of ZFL cells to model toxicants
Because CYP1A1 monooxygenase activity depends on AhR activation, we used the CYP1A1 promoter/enhancer region to design an AhR-responsive reporter construct. Previously, our study indicated that the 5' regulatory region is sufficient for transcriptional activation of CYP1A1 by AhR [26]. Before injecting zebrafish eggs with an EGFP reporter construct under regulatory control of the human CYP1A1 promoter, we had to determine whether this 5' regulatory region of CYP1A1 was responsive to AhR ligands in zebrafish. To test this, we constructed a plasmid by ligating the human CYP1A1 regulatory region to the cDNA sequence that encoded jellyfish GFP (Fig. 2). This construct was transiently transfected into adult ZFL cells. Cells were exposed to either DMSO (0.01%) or each toxicant, such as PCB126, TCDD, and 3-MC. Induction of EGFP activity was concentration-dependent in all groups (Fig. 3).
Discussion
The use of the zebrafish as an alternative model vertebrate for toxicology and pharmacology has only recently been initiated in Korea. The zebrafish model may provide useful information for recognizing and understanding the effects of in utero exposure to PCB126, TCDD, and 3-MC in humans. We determined developmental toxicities in early stages of developmental vertebrates with environtoxicants. One of the most striking responses to PCB126, TCDD, and 3-MC in zebrafish embryos was the accumulation of edematous fluid in the pericardium and the yolk sac. Edema was first observed in the pericardial region and yolk sac. Elongation and failure of the heart to undergo looping was also reported, and tail malformation was observed as teratogenic category. Gross malformations resulting from exposure to AhR-binding toxicants included jaw reductions, presumptive skeletal defects, and edema. Previously, certain classes of environmental contaminants were evaluated for early life stage toxicity in zebrafish [13,27]. In the present study, heart malformation was a characteristic feature following exposure to all three toxicants, and this may reflect the fact that edema can accompany cardiovascular dysfunction because the osmoregulatory function of the skin and the circulatory function of the heart and vasculature are correlated [5]. Moreover, zebrafish exposed to 100 nM PCB126 exhibited contorted tail and other tail malformation. This is the first report of PCB126 toxicity in zebrafish. The malformations were caused by the irregular blood flow of the caudal vein and artery following PCB126 exposure. Polychlorinated biphenyls (PCBs) have been analyzed extensively in various environmental samples since they were first identified and found to be very persistent [15]. PCB congener PCB 126 is the most potent congener, with a dioxin equivalency of approximately 0.l, compared to 1 by definition for TCDD [1,3,25]. The dioxin equivalency was approximately 0.07 (PCB126 EC50/TCDD EC50) in our results, which was also consistent with previous other reports.
PCB126, and TCDD bioaccumulated in fish that were exposed to these toxic substances [8,12,19]. PCB126 contributes most of the non-o-PCB dioxin-equivalents [24]. Bioconcentration factors are calculated on a wet weight basis ranging from 7,710 to 940,000 in zebrafish [7]. Indeed, the maximum contaminants level (MCL) of PCBs in drinking water is 5 × 10-4 mg/l, 1.53 × 10-9 M [29]. Thus, 1.53 × 10-9 M of PCBs in a body of water is concentrated 940,000 times to approximately 1.44 × 10-3 M PCBs in a fish, where it would act upon the AhRE motif. Nebert et al. [22] also emphasized the property of bioconcentration of environmental pollutants; for example, 10-17 M TCDD in a body of water is concentrated 100,000 times [8] to approximately 10-12 M TCDD is a fish, where it would act upon the AHRE motif [22]. Also, MCL of TCDD in drinking water is 3 × 10-8 mg/l, 0.93 × 10-13 M [25]. Thus, 0.93 × 10-13 M of TCDD in a body of water is concentrated 100,000 times [8] to approximately 0.93 × 10-8 M TCDD in a fish.
On the other hand, the transfected zebrafish cell line was validated using three toxicants that upregulated EGFP expression in a concentration-dependent manner, and therefore provided a reproducible in vitro tool for toxicological screening. The AhR signal transduction pathway in fish is similar to that in mammals [11,28]. Further studies on the mechanism of AhR ligands toxicity in a vertebrate model, the zebrafish, are warranted to better understand the effects of in utero AhR ligand-exposure in humans.
In the present study, we demonstrated that zebrafish might ultimately provide greater insight into the developmental toxicology of chemicals, as well as aid in low-cost, high-throughput screening in drug discovery and in evaluating the safety of large numbers of chemicals and nanomaterials [5].



 XML Download
XML Download