

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Antioxidants protect against radiation-induced oncogenic transformation in experimental systems [9]. Many natural and synthetic compounds have been investigated for their efficacy to protect against irradiation damage [35]. Previous studies developed radio-protective and radio-recovery agents to protect from the indirect effects of radiation by eliminating free radicals produced in response to radiation [54] and immunostimulants to counteract immune suppression [61]. Supplementary phytochemicals, including polyphynols, flavonoids, sulfhydryl compounds, plant extracts and immunomodulators, are antioxidants and radioprotective in experimental systems [55]. A potential treatment strategy for radiation exposure might be to strengthen the immune system [19].
The recent use of numerous herbal products as dietary supplements places them in a unique category of food to drugs (nutraceuticals) that are used for their therapeutic value. The realistic distinction between foods, dietary supplements, and drugs is often based on their future uses [10].
Echinacea (E.) purpurea was used to treat dizziness, snake bites and as an anti-infective agent until the advent of modern antibiotics [27]. Its recent resurgence as a treatment for recurrent genital herpes [57] and acute upper respiratory tract infections [46] has placed Echinacea among the most widely used herbs in the United States and Europe. In addition, Echinacea is also used as a preoperative herbal remedy [2], and it has anti-tumor [7] and anti-inflammatory [41] activities.
E. purpurea contains large amounts of chicoric acid and caftaric acid, which are largely recognized in the inhibition of hyaluronidase which is secreted by streptococci and other bacteria to enable penetration into tissue, has been demonstrated with Echinacea plant juice [31]. It also controls candidiasis infestation [22], enhances resistance to influenza viruses [49] and vesicular stomatitis virus [6] and enhances phagocytosis when administered orally to mice [12] and humans [26]. This phagocytic enhancement is attributed to its isobutylamide content, which inhibits the pro-inflammatory metabolite production induced by lipoxygenase [22] and is responsible for the local anaesthetic effects applied to relieve oral pain, such as toothaches and sore throats [37].
The "immune stimulation" by E. purpurea observed in-vitro and after parenteral administration has not been confirmed after oral intake in rats [52] and humans [47], and its preparations were immunologically inactive, even though they did show antioxidant and anti-inflammatory activities [42]. Other studies concerning the management of sinusitis in adults have demonstrated the efficacy of E. purpurea in the stimulation of the immune system, thereby reducing the incidence, duration and severity of respiratory infections [56]. The efficacy of E. purpurea has also been demonstrated in supportive treatment of urinary infections and for the external treatment of wounds and chronic arthritis [8]. New investigations have also shown that macrophage stimulation and the induction of cytokines are major parts of the mode of action [5]. Additionally, root extracts of E. purpurea were found to contain anti-oxidant compounds [39], to be capable of scavenging hydroxyl radicals and to suppress the oxidation of human low-density lipoprotein [21].
Most of the E. purpurea-related studies did not involve a thorough structural exploration of tissue proteins, particularly at the molecular level. Therefore, we used near-infrared Fourier transform Raman spectroscopy to study the structural changes of major liver constituents in irradiated mice.
The preventive and therapeutic properties of the immunomodulator and immunonutrient E. purpurea against radiation were reviewed by evaluating the changes in bone marrow and peripheral blood cell count and peripheral blood antioxidant activity.
Materials and Methods
Administration of E. purpurea
Standardized dried powder extract from E. purpurea (Echinacin; Madaus AG, Germany) at a dose of 30 mg/kg body wt/day, was suspended in 1.0 ml of saline and gavaged to each animal for 2 weeks as previously described by Di Carlo [13]. The dried powder extract from E. purpurea includes caffeic acid derivatives (primarily echinocoside), flavonoids, essential oils, polyacetylenes, alkylamides and polysaccharides [26].
Animals
Male Swiss albino mice aged 10 ± 1 weeks with an average weight of 21 ± 2 g were obtained from the Holding Company for Biological Products and Vaccines, Cairo, Egypt. The animals were kept under good ventilation, at a temperature of 22 ± 3℃, 60% humidity, and suitable illumination conditions (light/dark cycle of 14/10 h) and allowed maintenance nutrients and fresh water ad libitum.
Irradiation
A 137Cesium-γ-irradiator was provided by NCRRT, Egypt and was manufactured by Atomic Energy of Canada Ltd. The dose rate was 0.6 Gy/min of exposure.
Animal groups
A total of 120 mice were randomly divided into five groups of 24 animals each. In addition, each group was further divided according to the time of sacrifice.
Group A - Control group: Untreated and non-irradiated animals were given 1.0 ml normal saline/mouse/day for 2 weeks.
Group B - E. purpurea-treated group: Each mouse was given an appropriate dose of E. purpurea suspension/day for 2 weeks.
Group C - Irradiated group: Animals were subjected to one shot of whole body γ-rays (3 Gy) and then given 1.0 ml of normal saline/mouse/day for 2 weeks.
Group D - E. purpurea-treated and irradiated group (radio-protected group): Each mouse was given E. purpurea dosage/day for 2 weeks, and the animal was subjected to whole body γ-rays (3 Gy) at one hour after the last dose.
Group E - Irradiated and E. purpurea-treated group (radio-recovery group): Animals were subjected to whole body γ-rays (3 Gy), and each mouse was then given E. purpurea dosage/day for 2 weeks.
After the animals in the experimental group had been given all of the treatments, at intervals of 1, 2 & 4 weeks, the animals were sacrificed by cervical dislocation. Since leukocytes and erythrocytes have a relatively short life span of about 4 weeks in mice [54], the selected intervals were believed to reflect the hematological changes preceding irradiation [33].
Analytical methods
All hematological and biochemical parameters were performed according to standard laboratory methods using pure chemical materials from Sigma-Aldrich Co, USA.
Peripheral blood and bone marrow cell count
Peripheral blood samples were drawn from mice at experimental intervals of 1, 2 and 4 weeks. The hemoglobin (Hb) level, erythrocyte (RBC) count, total leukocyte (WBC) count and differential leukocytes (lymphocytes, neutrophils and monocytes) were investigated using an automated blood counter (Coulter Model T-450; Contronics, UK). On the same time intervals, femur bone marrow cells were prepared as described by Goldberg et al. [17]. Briefly, femoral bone was exposed under aseptic conditions, cells were washed with 199-medium (Sigma, USA), suspended by a syringe with a needles of various diameters, and washed again 2-3 times with 199 medium by repeated centrifugation at 150 × g for 10 min between each washing step. Smears of the cells were drawn on clean slides, fixed with methanol for 10 min and stained with May-Granwald-Giemsa (Sigma, USA). At least 1,000 cells were scored from each animal to determine the total myelokaryocyte count and differential elements (lymphoid & erythroid cells).
DNA fragmentation
Liver tissues (100 mg) were treated with 100 mM Tris-HCl, 5 mM EDTA, 150 mM sodium chloride and 0.5% sarkosyl, pH 8.0, at 4℃ for 10 min. Samples were incubated with ribonuclease (50 µg/ml) and proteinase K (100 µg/ml) for 2 h at 37℃ for 45 min. DNA was obtained by phenol:chloroform:isoamyl alcohol (25 : 24 : 1) (Sigma, USA) extraction, and precipitated with 0.3 M sodium chloride and cold isopropanol (1 : 1) at -20℃ for 12 h. DNA was recovered by centrifugation of the sample at 20,800 × g at 4℃ for 10 min. Thereafter, the precipitate was washed with 70% ethanol, dried and resuspended in Tris containing EDTA (10 mM Tris, 1 mM EDTA) at pH 8.0. Samples (100 µg DNA) were analyzed on a 1.5% agarose gel with ethidium bromide (0.5 µg/ml).
Raman measurement
Resonance Raman-spectroscopy was used as a fast and non-invasive optical method for measuring protein structural characterization in liver cells. FT-Raman with multiplex and high-throughput properties is able to obtain high-quality structural information at the molecular level. In the current study, near-infrared FT-Raman was used to detect the structural changes in the mouse liver following γ-irradiation and liver protection with E. purpurea.
FT-Raman spectra of liver tissues from the five groups were obtained using a Nicolet 670 spectrometer with the Nicolet Raman module 940 (Thermo Nicolet, USA) and Nd3+ laser operating at 1,064 nm with a maximum power of 2 W. The system was equipped with an InGaAs (Indium-Gallium Arsenide) detector, XT-KBr beam-splitter with 180-reflective optics, and a fully motorized sample position adjustment feature. A laser output power of 2 W was used and was low enough to prevent possible laser-induced sample damage and a high signal to noise ratio. Data were collected at 8 cm-1 resolution with 256 scans. Spectra were obtained in the Raman shift range between 400 and 3,700 cm-1. The system was operated using the OMNIC 5.3 software and the experiments were replicated three times. The intensity ratio of Raman bands 855-832 cm-1 (I855/832) was used to evaluate the microenvironment of tyrosine. Each numerical calculation of the Raman intensity ratio was based on the average of triplicate measurements.
Results
There were no significant differences between control groups and E. purpurea-treated groups in peripheral blood, bone marrow cell count, and peripheral blood antioxidant activities at any of the three time intervals. The time intervals of sacrificing also had no effect on the above-mentioned parameters within the five study groups (Tables 1-4).
As shown in Table 1, the Hb levels and RBC and WBC counts in the γ-irradiated groups showed significant decreases in comparison with the control and E. purpurea-treated groups. However, there were significant differences in Hb level at week 4 as compared with weeks 1 and 2.
In the E. purpurea-treated group followed by irradiation (radio-protected group) and irradiated group followed by E. purpurea-treatment (radio-recovery group), Hb levels during the three time intervals increased significantly in comparison to the control group, but the difference in Hb level in the radio-recovery group, E. purpurea treated group and that of the control and radio-protected groups became insignificant at week 4.
The RBC count in both the radio-protected and radio-recovery groups increased significantly in comparison with the irradiated group at each of the three time intervals. The RBC count tended to increase towards the counts in both the control group and E. purpurea-treated group at weeks 2 and 4, in both the radio-protected and radio-recovery groups. In addition, the total WBC count increased significantly in both the radio-protected and radio-recovery groups in comparison with the irradiated group at each of the three time intervals. There were significant differences between the total WBC count at the 4th week as compared with its level at the 1st week in the radio-protected group while the differences in total WBC count were significant in the radio-recovery groups at week 4 in comparison with its level during both the 1st and 2nd weeks (Table 1).
As shown in Table 2, the γ-irradiated groups showed significant decreases in the numbers of lymphocytes, neutrophils and monocytes as compared with the control group and E. purpurea-treated groups. In both the radio-protected and radio-recovery groups, the differential leukocyte counts tended to increase towards the normal levels at all three time intervals.
As shown in Table 3, the γ-irradiated groups showed marked and significant decreases in the numbers of myelokaryocyte, lymphoid and erythroid cells as compared with the control and E. purpurea-treated groups. The time intervals had effects on the numbers of myelokaryocyte and lymphoid cells at weeks 1 and 2 and in the three types of cells at all three time intervals. In both the radio-protected and radio-recovery groups, the counts of the three types of cells increased significantly compared to the irradiated group until the difference in monotype cells between the radio-protected and radio-recovery groups and the control and E. purpurea-treated groups became insignificant at the three time intervals (Table 3).
As shown in Table 4, the γ-irradiated groups showed marked and significant augmentation in TBARs levels at the three time intervals as compared with its level in both the control and E. purpurea-treated groups. There was a significant difference between the TBARs values at weeks 1 and 4 in the γ-irradiated groups. In contrast, there were significant decreases in SOD and GSPx activities as compared to the control and E. purpurea-treated groups. There were also significant differences in TBARs value at weeks 1 and 4 in the γ-irradiated groups.
The TBARs levels in both the radio-protected and radio-recovery groups decreased significantly as compared with the γ-irradiated groups, but the decrease in the radio-protected groups was greater than that in the radio-recovery groups at the three time intervals. The activities of SOD and GSP increased significantly in both the radio-protected and radio-recovery groups as compared with the γ-irradiated groups (Table 4).
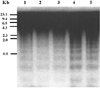
The administration of E. purpurea before γ-exposure reduced apoptosis as measured by DNA fragmentation (Fig. 1). In our experiments, the DNA fragmentation in the mouse liver cells could not be recovered by the administration of E. purpurea after γ-irradiation.
Fig. 2 shows the Raman spectra, which ranged from 400 to 3,700 cm-1, of mouse livers in the control group (A), E. purpurea-treated group (B), 3 Gy gamma-irradiated group (C), radio-protected group (D) and radio-recovery group (E). The secondary structure information, primarily seen as antiparallel β-pleated sheets, was indicated by the vibrational stretch of amide I (~1,670 cm-1) and amide III (~1,241 cm-1) only in Groups A and B. The secondary structure of the protein in the mouse liver was not stable in C and amide I was shifted to ~1,590 cm-1 while amide III was stable. In Group D, the secondary structure of the protein in liver cells was stable enough to resist changes in the spectra. The comparison of the Raman spectra of the radio-recovery group (E) in Fig. 2 and the control group (A) showed that the vibrational stretch of amide I was shifted to ~1,620 cm-1. The vibrational stretch of amide III in Group C could not be detected by Raman spectra, while the amide III in Group E was shifted to 1,180 cm-1.
The tyrosine residues were detected at the doublet Raman shift of 855 and 832 cm-1. The ratio of both doublets indicated the hydrogen bonding environment in the liver. The intensity ratios (I855/832) for A, B, C, D, and E Group were 0.48, 0.47, 1.25, 0.62 and 1.17, respectively. In other words, the tyrosine residues in Group C were greatly affected by radiation. The tyrosine residues in Group D were more susceptible to E. purpurea treatment before radiation than the E. purpurea treatment after radiation (Group E).
Discussion
E. purpurea has generally been considered to be safe and without significant toxicity, significant herb-drug interactions, contraindications, or adverse side effects [8,23, 28].
The hematopoietic system is known to be one of the most radiosensitive systems, and its damage may play lead to the development of hematopoietic syndrome and result in death. Survival after irradiation actually results from the recovery of several target systems, such as the bone marrow, gastrointestinal tract, skin and hemostatic systems [59]. Death from the so-called hematopoietic syndrome results from infection due to the impairment of the immune system [11]. Various mechanisms, such as the prevention of damage through the inhibition of free radical generation or its intensified scavenging, enhancement of DNA and membrane repair, replacement of dead hematopoietic and other cells and the stimulation of immune-cells activities, are considered to be important approaches for radio-protection and radio-recovery [36].
In the present study, the reduction in both Hb level and RBC count at each of the three time intervals in the irradiated groups were attributed to the impairment of cell division, obliteration of blood-forming organs, alimentary tract injury [14], depletion of factors needed for erythroblast differentiation and reticulocyte release from the bone marrow [18] and the loss of cells from the circulation by hemorrhage or leakage through capillary walls and/or the direct destruction of mature circulating cells [53]. Recovery of both Hb level and RBC count was evident in both the protected and recovered groups, but the recovery of the Hb parameter was more distinct in the radio-protection group than in the radio-recovery group. In contrast, the RBC counts in the radio-protection and radio-recovery groups were the same as those of the control, E. purpurea-treated and irradiated groups.
The present work describes the marked decrease in WBC count in mice subjected to irradiation at three time intervals. Irradiation-induced leucopoenia has likewise been reported in γ-ray irradiated mice [33]. It seems apparent that the leucopoenia observed in these mice was a direct consequence of the lymphopenia and neutropenia that occurred following irradiation. An obvious degree of either radio-protection or radio-recovery was obtained using E. purpurea. These results agree with the findings of Barrett [3] and Widel [59], who reported that Echinacea preparations influenced the leukocyte count, stimulated the phagocytic activity and/or increased the release of cytokines. It has been suggested that Echinacea is able to stimulate innate immune responses, including those regulated by macrophages and natural killer cells (white blood cells). In addition, macrophages respond to purified polysaccharide and alkylamide preparations incorporated into Echinacea. Treatment with ionizing radiation resulted in cytokine-mediated cellular damage [30]. For patients undergoing radiation and chemotherapy treatments, studies have proven that E. purpurea, while boosting the immune system, also produced additional white blood cells and stimulated bone marrow production, which was diminished by chemotherapy [43]. However, the mechanisms of stimulation for cells responsible for adaptive immunity have not been fully elucidated for the other molecules present in E. purpurea preparations [22].
Since the peripheral blood pattern observed during the entire post-irradiation period was primarily a reflection of processes occurring in hematopoietic organs [59], the significant protective effects of E. purpurea against lymphoid cell death in bone marrow can lead to their accelerated recovery in peripheral blood. In fact, the tendency to return to the normal value of reduced blood leukocyte count throughout the three time intervals was more rapid in the E. purpurea-treated groups both before and after irradiation than in the irradiated mice.
In this study, lymphocytes, neutrophils and monocytes were significantly decreased throughout the three time intervals. Mature lymphocytes are considered to be the most sensitive type of blood cell [60], and the earliest blood change following whole body irradiation is lymphopenia [45]. Neutrophils have a half-life of only about 10-12 h once they leave the marrow, a site that serves as a reservoir for mature neutrophils [34]. These data agree with the findings of Kafafy et al. [25]. The data showed that E. purpurea administration has significant radio-protective and radio-recovery effects on the levels of lymphocytes, neutrophils and monocytes.
It has been reported that E. purpurea has an IFN-like effect, activating macrophages and inducing the production of interleukin -1 (IL-1) and IFN [48]. In addition, Mishima et al. [33] reported that the administration of E. purpurea had a suppressive effect on radiation-induced leucopoenia, especially on lymphocytes and monocytes, and resulted in a faster recovery of the blood cell count in mice and rabbits [24]. In addition, peripheral blood antioxidant activity was increased by E. purpurea, which suggested a relationship between the antioxidant effect and the suppressive effects on radiation-induced leucopoenia. In contrast, Schwarz et al. [48] reported that the oral administration of E. purpurea for 2 weeks had only minor effects on 2 out of 12 lymphocyte subpopulations determined by flow-cytometry in a double-blind, placebo-controlled cross-over study.
In the present study, irradiation caused remarkable increases in the TBARs content and incredible decreases in the activities of SOD and GSPx. Zahran et al. [63] and Tawfik et al. [54] recently confirmed these finding. After the administration of E. purpurea, the TBARs level and antioxidant activities were attenuated in comparison to their values in irradiated mice at each of the three time intervals. The mechanisms of antioxidant activity in the extracts derived from Echinacea included free radical scavenging and transition metal chelating properties [23].
Several experimental models have described the in vivo and in vitro protection from liver injury induced by free radicals [1,40]. They reported that prostaglandin (PGE1) was able to reduce DNA fragmentation in rat hepatocytes and that it protected against galactosamine (D-GalN)-induced apoptosis. It is interesting to note that the administration of Echinacea also reduced the effects of gamma irradiation on DNA fragmentation. In contrast, the administration of Echinacea after gamma exposure was not effective at reducing the apoptotic mechanisms induced by gamma irradiation. The protection provided by Echinacea against apoptosis induced by gamma irradiation may be associated with its ability to block the induction of internal factors, such as inducible nitric oxide synthase (iNOS) and nitric oxide (NO) production. In fact, Echinacea was able to slightly enhance DNA fragmentation in control cells. Nevertheless, more studies are needed in order to confirm these findings.
The secondary structure of liver proteins is easily monitored by observing the frequencies of amide I and amide III originating from a peptide backbone [50]. The sharpening of amide I peaks in A, B and D may indicate the uniformity of hydrogen bonds whereas the flattening of amide I in the gamma-irradiated mice (Group C) and its shifting to 1,590 cm-1 may indicate the loss of uniformity in hydrogen bonds.
In the liver, tyrosine is a key component of many enzymes, which may be inhibited through the oxidative modification of their tyrosine residues. Therefore, it is very important to probe the microenvironment of tyrosine. Shih et al. [51] reported that the tyrosine doublet at 850-1 and 830 cm-1 was sensitive to the nature of the hydrogen bond of the phenol hydroxyl group. If a tyrosine residue is on the surface of a protein in aqueous solution, the phenolic OH will simultaneously act as an acceptor and donor of moderate to weak H-bonds, and the doublet intensity ratio (I850/830) will be about 1 : 0.8 (I = 1 : 25). If the phenolic oxygen is the acceptor atom in a strong H-bond, the intensity ratio will be about 1 : 0.4 (I = 2 : 5). If the phenolic hydroxyl is the proton donor in a strong H-bond, the intensity ratio will be approximately 1 : 2 (I = 0.5). Accordingly, the current result that the intensity ratio in the gamma-irradiated mice (Group C) was about 1 : 0.8 might indicate that the phenolic hydroxyl of tyrosine was on the surface of liver proteins with a moderate to weak H-bond. On the other hand, the doublet intensity ratio in the radio-protected mice (Group D) was sensitive to the level of E. purpurea administration, as shown in Fig. 2b. However, the mechanisms by which these antioxidative effects protect major liver constituents, including thiol compounds, tyrosine, tryptophan, and water content, from oxidative insults remains to be elucidated.
Weiss and Landauer [58] documented a protective effect of polyphenols from Echinacea against free radical damage and a class of specific antioxidants known as caffeoyl derivatives in appreciable amounts. Furthermore, Sasagawa et al. [44] reported that the alkylamides present in Echinacea species inhibited IL action and hypothesized that the constituents present in its dry extracts exert direct immunomodulatory effects on the immune system [44]. In addition, single X-ray irradiation causes considerable disturbances to the liver. The administration of Echinacea tinctures was assumed to induce their beneficial effects, primarily by stimulating certain components of the non-specific immune system. Previous studies have proven that the most important pharmacological effects were the stimulation of the phagocytic activity of polymorphonuclear leucocytes and other phagocytes [3], as well as the activation of phagocytes to produce the pro-inflammatory cytokines TNF-α, IL-1, IL-6 and other mediators [4].
E. purpurea was able to regulate the process of apoptosis in-vivo. The splenic-lymphocytes from mice orally treated with Echinacea for 14 days at a dose of level 30 mg kg-1 per day were shown to be significantly more resistant to apoptosis than those from mice treated only with the vehicle [13]. Moreover, Gan et al. [16] demonstrated that Echinacea extracts are potent activators of natural killer (NK) cytotoxicity, augmented the frequency of NK target conjugates and activated the programming for NK cell lysis. The Echinacea extracts also enhanced the antibody-forming cell response and humeral immune responses as well as the innate immune responses in female mice [15]. It also enhanced the nonspecific immune or cellular immune systems (or both) in the AKR/J-mice [20]. It also sensitized the immune cells and led to lifespan prolongation in mice [12].
Raso et al. [41] evaluated the anti-inflammatory activity of E. purpurea in mice treated at doses of 30 and 100 mg kg-1 twice daily. Only the higher-dose treatment significantly inhibited the formation of edema in a time-dependent manner. Western blot analysis showed that in vivo treatment with this extract could modulate lipo-polysaccharide and INF-γ-induced cyclooxygenase-2 (COX-2) and iNOS expression in peritoneal macrophages. They suggested that the anti-inflammatory effect of that particular extract could be in part related to its modulation of COX-2 expression.
The mechanisms of the stimulatory effect observed in the present study remain to be clarified. The authors suggest that the factors that might be involved are changes in the intestinal absorption of immune stimulating-compounds present in the Echinacea preparation caused by the irradiation. Brinker [10] reported that the experimental success of the oral administration of the immunostimulant E. purpurea was probably due to the receptor binding of its polymeric markers on mucosal- or gut-associated lymphoid tissues.
In conclusion, the immune stimulatory ability of E. purpurea extracts may have a therapeutic potential to regulate the protection and recovery of immune responses as well as the activation measures in irradiated mice.
Therefore, further studies are needed to clarify the mechanism(s) that are responsible for the beneficial effect of Echinacea preparations observed in this study, and future research must also be conducted on the use of E. purpurea as an immunonutrient and useful adjunct to conventional cancer therapies because of its immune-stimulating properties.





 XML Download
XML Download