

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Dengue virus infection is the most common arthropod-borne viral infection worldwide (1). Most instances of dengue virus infection are asymptomatic or result in a short-lasting undifferentiated febrile illness, especially in children (2). Symptomatic dengue virus infection often results in a characteristic self-limited illness described as classical dengue fever (DF). Dengue haemorrhagic fever (DHF) is an infrequent but potentially lethal form of illness that develops usually following secondary infection with a heterologous serotype of dengue virus (1). The hallmark of DHF is the occurrence of plasma leakage (3). Although it is widely accepted that immunopathology underlies the development of plasma leakage, the pathogenesis of DHF is incompletely understood (3). Higher plasma viraemia early in the course of infection and consequent T-cell activation are known to predispose to the development of DHF (4,5). Macrophage inflammatory protein-1α (MIP-1α) is a chemokine that plays an important role in the immunopathogenesis of viral infections. Recent in vitro studies have suggested a pathogenic role for MIP-1α in dengue virus infection as well (6).
Earlier, we have reported the occurrence of a TH2-skewed immune response around the time of defervescence in a group of patients comprising mostly of those with DF (7). We noted a trend towards a higher intracellular interferon-γ (IFN-γ)/interleukin-4 (IL-4) ratio and lower serum IgE levels in patients with DHF as compared to those with DF. Based on these preliminary findings, we postulated that an unopposed TH1 type immune response contributes to the pathogenesis of DHF (7). In the present study, we tested this hypothesis in a larger group of patients with varying severity of dengue. In addition, we also attempted to define the role of MIP-1α as an immunologic correlate of disease severity in patients with dengue.
MATERIALS AND METHODS
Subjects
This study was conducted at the All India Institute of Medical Sciences hospital, New Delhi, India. This is a large tertiary level teaching hospital located in northern India. In this part of the world dengue transmission is largely seasonal, punctuated by several epidemics in the past, the most recent one being in the year 2003. An outbreak of dengue occurred in New Delhi during the months of October-November 2006 resulting in about 3,300 cases and 65 deaths (8). We prospectively enrolled patients aged more than 12 years with serologically confirmed dengue virus infection hospitalised during this outbreak. In order to circumvent the uncertainty regarding the classification of disease severity using the current World Health Organization (WHO) definitions (9) and to elucidate clearly the differences in immune response between patients with DF and DHF, we employed a purposive sampling strategy - patients with DHF recruited in the study had unequivocal features of plasma leakage evidenced by the presence of haemoconcentration along with serosal effusion(s), and those in the DF group had none of these features. We graded the severity of DHF as per the WHO classification (10). None of the patients were atopic or reported any risk factor for human immunodeficiency virus infection. We also screened 20 healthy, non-atopic volunteers with no history of DF/DHF in the past for the presence of dengue-specific serum IgG and IgM antibodies. After screening, we chose 10 dengue-naive volunteers (five males) as controls for immunological studies. The study was conducted in compliance with the ethical guidelines for biomedical research on human subjects, Indian Council of Medical Research, New Delhi, India. We obtained informed consent from all study patients/volunteers or their legal guardians.
We prospectively collected data on demographic characteristics, clinical findings, and laboratory findings such as serial haematocrit and platelet counts, total and differential leukocyte counts, and liver function tests using a predesigned instrument. Dengue viral RNA detection and serotype identification was done by a reverse transcriptase polymerase chain reaction (RT-PCR) assay, as described earlier (11). Dengue-specific IgM and IgG antibodies in the serum were estimated using commercially available capture enzyme-linked immunosorbent assay kits (Panbio Ltd., Queensland, Australia). After initial fluid resuscitation and stabilization, we collected peripheral venous blood specimens for immunological studies (baseline specimen), and repeated the immunological studies after 48 h and on day 5 of hospitalisation. In order to account for the diurnal rhythm in TH1-TH2 balance, we collected the blood specimens for immunological studies in the morning at approximately the same time within a time window of 2 h.
Immunological studies
Peripheral blood mononuclear cells (PBMCs) were isolated by Ficoll-Hypaque density gradient centrifugation, washed twice, and resuspended in RPMI 1640 cell culture medium supplemented with 10% inactivated foetal bovine serum. PBMCs were stimulated with phorbol myristate acetate (1 ng/ml) and ionomycin (1µM) in the presence of the Golgi transport inhibitor, monensin (2µM), for 6 h in a humidified 5% CO2 incubator at 37℃. Following stimulation, the PBMCs were collected, and immunostaining was done as per the manufacturer's specifications (BD Pharmingen, BD Biosciences, San Diego, CA). Surface staining for CD8 was performed with Cy5PE-labelled monoclonal antibodies at 4℃ for 15 min. For intracellular immunostaining, stimulated PBMCs were fixed in 2% formaldehyde and permeabilised with 0.5% saponin, followed by staining with fluorescein isothiocyanate-labelled IL-4 and tumor necrosis factor-α (TNF-α) monoclonal antibodies and phycoerythrin-labelled IFN-γ and MIP-1α monoclonal antibodies. Samples were run in a flow cytometer (FACSCalibur, BD Biosciences, CA) within 24 h of staining and analysed using FlowJo cytometry analysis software. Serum samples were stored at -20℃. Estimation of serum IFN-γ, IL-4, and TNF-α was performed in duplicate using commercially available enzyme-linked immunosorbent assay kits (BD Biosciences, San Diego, CA). The lower limit of detection for these assays was 2 pg/ml.
Statistical analysis
We performed all statistical analyses using a software package (SPSS for Windows, version 11.5, SPSS Inc., Chicago, IL). We present normally distributed continuous variables as mean±SD and compared them between the DF and DHF groups using independent t-test. We present continuous variables with a skewed distribution as median (interquartile range) and compared them between the groups using Mann-Whitney U test. We compared serial observations within a group using a nonparametric equivalent of repeated measures ANOVA (Friedman test), and applied the Spearman's rank correlation to test the correlation between two continuous variables. We applied the Cuzick test to test for a linear trend across the ordered groups, namely, controls vs. DF vs. DHF. We calculated percentage-change in haematocrit as [(haematocrithighest-haematocritdischarge)/haematocritdischarge]. We present the categorical variables as number (%) of patients and compared them between groups by chi-squared test. In addition, we carried out stratified analyses to evaluate the effect of the following baseline characteristics - gender, duration of fever at presentation, and dengue virus serotype - on the IFN-γ/IL-4 ratio in CD4+ T-cells. All tests were two-sided, and p<0.05 was considered statistically significant.
RESULTS
We studied 20 patients (DF=10, DHF=10) and 10 dengue-naïve healthy controls. Mean age of the patients and controls was 29±9 and 23±3 years, respectively. Among the patients, the median duration of fever at the time of hospitalisation was 5 (3~7) days. Of the 10 patients with DHF, four had grade I, five had grade II, and the remaining one patient had grade III DHF. All patients except the one that had grade III DHF recovered from the illness uneventfully and survived to hospital discharge. Based on the dengue-specific antibody responses in the serum, all patients except one with DHF were characterized as secondary dengue virus infections. Dengue viral RNA was detected by RT-PCR in 10 of 20 patients tested - serotype 3 (DENV3) in four and serotype 4 (DENV4) in six patients respectively; of these 10 patients, eight had DF (four each of DENV3 and DENV4) and the remaining two patients with DENV4 infection had DHF. Age, duration of fever at presentation, and platelet count at admission were comparable between patients in the DF and DHF groups (Table I). All patients in the DHF group were male as compared to the DF group in which males comprised only about a half of the group. Serum alanine aminotransferase (ALT) levels were significantly higher in the DHF group (Table I).
As compared to dengue-naïve controls, the proportion of IFN-γ-positive CD4+ T-cells as well as CD8+ T-cells was markedly higher at baseline in both DF and DHF groups, and an increasing trend was observed across the three groups namely, controls vs. DF vs. DHF (p<0.001 for both trends; Table II, Fig. 1). Similarly, an increasing trend across these groups was also noted in the proportion of MIP-1α-positive CD8+ as well as CD4+ T-cells (p<0.001 for both) and the IFN-γ/IL-4 ratio in the CD4+ as well as the CD8+ T-cell compartments (p<0.001 and p=0.004, respectively; Fig. 2). Interestingly, IL-4-positive CD4+ T-cells were significantly higher in the DF group (p=0.023), but not in the DHF group (p=0.912), as compared to controls (Table II).
As compared to DF, in patients with DHF the proportion of CD4+ T-cells positive for intracellular IFN-γ was higher at baseline and those positive for IL-4 was lower (Table II), resulting in a significantly higher IFN-γ/IL-4 ratio in the CD4 + T-cell compartment [7.21 (5.36~10.81) vs. 3.04 (1.75~4.02); Fig. 1; p=0.011]. Similarly, proportion of MIP-1α-positive CD8+ T-cells was significantly higher in the DHF group (Table II; p=0.023). Further, there was a strong positive correlation between IFN-γ/IL-4 ratio in the CD4+ T-cell compartment and MIP-1α in CD8+ T-cells (Fig. 3A; Spearman's rho=0.64; p=0.003). Proportion of MIP-1α-positive CD8+ T-cells also had a significant positive correlation with the percentage-change in the haematocrit (Fig. 3B; Spearman's rho=0.47; p=0.048) and with serum levels of ALT (Fig. 3C; Spearman's rho=0.61; p=0.009). However, it did not correlate with platelet count nadir, absolute neutrophil count, and total lymphocyte count (data not shown).
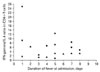
On stratified analysis, when the analysis was restricted to males, IFN-γ/IL-4 ratio in the CD4+ T-cell compartment remained comparatively higher in patients with DHF [7.21 (5.36~10.81) vs. 2.86 (2.00~5.55)]. When individually plotted according to the duration of fever at presentation, the IFN-γ/IL-4 ratio in CD4+ T-cells was higher in all patients with DHF except one as compared to patients with DF of the same duration of fever (Fig. 4). The IFN-γ/IL-4 ratio in the CD4+ T-cell compartment was similar between patients with DENV3 and DENV4 infections in the DF group [2.87 (1.80~5.58) vs. 2.36 (1.61~5.47)].
On serial estimation (Table II), in patients with DF, there was a significant decline in the proportion of IFN-γ-positive CD8+ T-cells (p=0.021) and a trend towards an increase in TNF-α-positive CD8+ T-cells (p=0.066). Further, in this group, a declining trend was observed in the proportion of IL-4-positive CD4+ T-cells (p=0.066), while IFN-γ-positive CD4+ T-cells and MIP-1α-positive CD8+ T-cells showed no significant change over time. In patients with DHF, there was a significant decline in the proportion of IFN-γ-positive CD8+ T-cells (p=0.005), as observed in the other group (Fig. 4).
In the serum, at baseline, measurable quantities of IL-4 were found in 4 of 10 patients in the DF group, while it was undetectable despite repeated estimations in all patients with DHF. Importantly, a conspicuous discordance between serum and intracellular IFN-γ was noted in patients with DHF - serum IFN-γ levels were comparatively lower in patients with DHF as compared to DF at all time points of the study despite higher IFN-γ in the T-cell compartments (Table II). A declining trend on serial estimation was observed in the serum levels of TNF-α in patients with DHF (p=0.075).
DISCUSSION
We found that the intracellular IFN-γ/IL-4 ratio in CD4+ T-cells was significantly higher in patients with DHF, suggesting a TH1-skewed immune response in these patients. The present study extends our earlier findings and confirms the hypothesis that DHF is associated with a TH1-skewed immune response around the time of defervescence vis a vis a TH2-skewed response in patients with DF. We also found that MIP-1α in CD8+ T-cells positively correlated with certain important markers of disease severity such as percentage-change in haematocrit and serum ALT levels. These markers are directly reflective of two important events in the pathogenesis of DHF namely vascular leakage and hepatocyte injury, respectively (1,12). MIP-1α is a CC chemokine with pro-inflammatory and stem cell inhibitory activities in vitro and is known to mediate virus-induced inflammation in vivo (13). Correlation of MIP-1α with the markers of disease severity suggests that the former might be an important mediator of immunopathology in DHF. To the best of our knowledge, the association between MIP-1α and disease severity in patients with dengue has not been reported earlier, although it has been hypothesised based on in vitro findings (6). It has also been suggested that MIP-1α might be responsible for the transient bone marrow suppression seen in patients with dengue (14). We, however, did not observe any significant association between MIP-1α and the parameters reflective of bone marrow suppression in the present study.
There are at least four potential confounding variables that could account for a higher IFN-γ/IL-4 ratio in patients with DHF - i) primary vs. secondary immune response; ii) duration of fever at presentation; iii) gender; and iv) differences in infecting serotype. Serendipitously, all but one patients included in the present study had secondary dengue virus infection. This negates confounding by the first of these four variables, and stratified analyses do not suggest the presence of confounding by any of the remaining three variables. Thus, we consider that the observed difference was in fact attributable to difference in disease severity.
There are at least two potential explanations for this observed association of a TH1-skewed immune response with DHF. First, antibody-dependent enhancement of dengue virus infection is a key mechanism thought to underlie the pathogenesis of DHF (15). IFN-γ is known to enhance the Fcγ receptor-mediated infection of monocytic cells by the dengue virus bound to cross-reactive antibodies (16); elevated levels of IFN-γ production by TH1-polarised T-cells would thus result in an increased infected cell mass. Second, TH1-polarised cytokine(s) enhances presentation of viral epitopes by professional antigen presenting cells to CD8+ T-cells, leading to their activation (17). CD8+ cytotoxic T-cells play an important role in viral clearance and immunopathology of dengue virus infection (18,19).
On the other hand, activated CD8+ T-cells are a biologically important source of MIP-1α in intracellular infection models (20), and MIP-1α is known to skew the immune response towards a TH1 polarity (21). Thus, the positive correlation between IFN-γ/IL-4 ratio in CD4+ T-cells and MIP-1α in CD8+ T-cells, observed in the current study, is probably the result of a bidirectional positive feedback loop operating between these two compartments. Earlier studies have established the association of early immune activation (in quantitative terms) with the subsequent development of DHF in patients with dengue virus infection (Green S et al., 1999). In the present study, we have characterised this immune response as a TH1-skewed response at the intracellular level. In a well-conducted study by Green et al. (5), it was observed that the plasma IFN-γ levels abruptly increased in children with DHF just preceding the defervescence of fever followed by a decline. The findings of the present study suggest the occurrence of a similar phenomenon at the intracellular level as well. The current understanding of the pathogenesis of DHF is far from complete and controversial. Though an oversimplification, it is generally agreed that the positive feedback of IFN-γ secreted by activated T-cells on macrophages sets off a vicious cycle of dysregulated cytokine production leading to DHF. A TH1-skewed immune response in patients with DHF as found in the present study is in consonance with this concept.
Notwithstanding, another school of thought holds that DHF is associated with a TH2 type immune response (22,23). There are several important limitations that could have biased the findings of these studies - first, the patients were classified as DF or DHF based on assessment at the time of enrollment (24). Subsequent clinical progression to DHF of patients initially classified as DF, which is a common phenomenon, was not taken into account, and this could have resulted in misclassification of cases. Second, the uncertainty in classification of disease severity using the WHO definitions (9) could also have contributed to misclassification of patients to an unknown extent in these studies. We circumvented these biases by adopting a purposive sampling strategy. Third, the increased vascular permeability in DHF is likely to cause a leakage of secreted cytokines into the extravascular space resulting in a discordance of serum levels with the actual intracellular expression of cytokines - we found suggestive evidence for the existence of such discordance in the present study. If one estimates only the serum levels of cytokines (24), it would paradoxically appear that the very same cytokines associated with DHF are comparatively lower in the sera of patients with DHF as compared to DF. This could explain the fact why earlier studies that estimated cytokine levels only in the serum of patients with DHF found a TH2-skewed pattern even though at the intracellular level the T-cell response is actually TH1-skewed in nature.
Cell-mediated immunity is a very important mechanism by which the host clears off an established viral infection. An exuberant cell-mediated immune response also clears the viral infection albeit at the cost of considerable immunopathology. Despite the lack of any specific anti-viral treatment, the mortality in patients with DHF has been brought down to less than 1% with supportive management alone (25); this implies that the immune response alone manages to successfully clear off the viraemia in all these patients. If DHF is characterised by a TH2-skewed immune response, as proposed by some authors, it cannot explain this observation. Further ancillary evidence to our finding that DHF is characterised by a TH1-skewed immune response comes from the epidemiologic observation that malnutrition decreases the risk of DHF in patients with dengue virus infection (26). Malnutrition is known to impair the cell-mediated immune response resulting in a shift towards a TH2 profile (27). Though the present study brings out the association between a TH1-skewed immune response and DHF, it does not tell us exactly which factor tipped off the balance towards a TH1-skewed CD4+ T-cell response. We believe the missing link could be the expansion of regulatory T-cells in patients with milder disease which suppresses the IFN-γ production (28) possibly skewing the immune response towards a TH2 profile in DF.
To conclude, we found that the intracellular IFN-γ/IL-4 ratio in CD4+ T-cells and MIP-1α in CD8+ T-cells were significantly higher in patients with DHF. We also observed that MIP-1α in CD8+ T-cells positively correlated with the intracellular IFN-γ/IL-4 ratio in CD4+ T-cells as well as important markers of disease severity in patients with dengue virus infection. These findings confirm our hypothesis that an unopposed TH1 immune response underlies the pathogenesis of DHF. The present findings also suggest that MIP-1α might be an important mediator of immunopathology in DHF.





 XML Download
XML Download