

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Carbapenems such as imipenem and meropenem are first-line drugs in the treatment of serious infections caused by multidrug-resistant Pseudomonas aeruginosa, Acinetobacter spp., and Enterobacteriaceae. However, in recent years, imipenem resistance in P. aeruginosa and Acinetobacter spp. has been increasing steadily around the world, and in Korea, these resistance rates reached 22% and 64%, respectively, in 2011.1 Among several mechanisms of carbapenem resistance, acquired metallo-β-lactamases (MBLs) have a more serious impact as the enzymes confer a high level of resistance and the genes can be transferred horizontally.2,3 Among acquired MBLs, VIM-type and IMP-type enzymes are the most common types of MBLs with worldwide distribution,4 and the VIM-2 type has been highly prevalent in Korea.5,6 In our previous study in 2003-2004, imipenem-nonsusceptible P. aeruginosa isolates carrying the blaVIM-2 allele were highly prevalent, and the incidence of Acinetobacter spp. carrying the blaVIM-2 allele had increased compared to those carrying the blaIMP-1 allele.5 The aim of the present study was to determine the trends of carbapenem-resistant and MBL-producing Gram-negative bacilli over the past 8 years in a Korean teaching hospital with more than 2000 beds. This is a unique report on the long term trend of carbapenem-resistant and MBL-producing Gram-negative bacilli isolated in a single hospital.
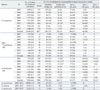
In total, non-duplicated clinical isolates of 12650 P. aeruginosa, 1096 other Pseudomonas spp., and 7650 Acinetobacter spp. in addition to 14026 Klebsiella pneumoniae, 6110 Enterobacter cloacae, and 3162 Serratia marcescens isolates among Enterobacteriaceae were recovered from patients at the hospital from 2005 to 2012 (Table 1). The isolates were identified by conventional methods using ATB 32 GN or VITEK-2 systems (bioMerieux, Marcy-l'E'toile, France). Antimicrobial susceptibilities were determined using the disk-diffusion method or the VITEK-2 system (bioMerieux). The Clinical and Laboratory Standards Institute 2010 breakpoints were used after January 2011.7 The modified Hodge test (MHT) and the imipenem and ethylenediaminetetraacetic acid sodium mercaptoacetic acid double disk potentiation (IEDDP) test were conducted to screen for MBL producers in imipenem- or meropenem-nonsusceptible isolates.8 Polymerase chain reaction (PCR) was performed to detect and sequence blaVIM-2-like, blaIMP-1-like and blaSIM-1-like genes.9,10 In carbapenem-nonsusceptible strains showing MHT-positive yet the aforementioned PCR negative results, blaNDM and blaKPC genes were also screened by PCR.11,12 The nucleotide sequences of PCR-generated amplicons were analyzed for the representative strains. XbaI- or SmaI-digested genomic DNAs from 75 P. aeruginosa, 8 P. putida, and 109 Acinetobacter spp., all of which were randomly selected MBL-producing spp. isolated in 2005-2006, were separated by pulsed field gel electrophoresis (PFGE) using a CHEF-DR II system (Bio-Rad, Hercules, CA, USA) and BioNumerics software v. 5.10 (Applied Maths, Sint-Martens-Latem, Belgium). PFGE banding pattern clustering with an 80% similarity threshold was determined using the Dice coefficients and the unweighted pair group method using arithmetic averages using Molecular Analyst Fingerprinting Software (Bio-Rad). Related clones with one or two independent genetic events were designated as subtype numbers in Table 2. The S1-digested DNA of randomly selected P. aeruginosa, Pseudomonas putida, and Acinetobacter spp. strains carrying the blaVIM-2 allele were blotted onto nylon membranes (Bio-Rad) and hybridized with blaVIM-2 gene probes to observe the differences in the plasmids carrying the gene.
While annual imipenem resistance rates in P. aeruginosa increased from 14% in 2003 to 29% in 2009, they decreased from 58% in 2003 to 33% in 2012 in other Pseudomonas spp.; in Acinetobacter spp., resistance rates increased steeply from 13% in 2003 to 66% in 2012.5 However, the imipenem resistance rates in K. pneumoniae, E. cloacae, and S. marcescens isolates remained at less than 1%. The positive rates of MHT were highly variable depending on the species, being only 13% in P. aeruginosa isolates, yet reaching 69% in other Pseudomonas spp. and 94% in Acinetobacter spp. These findings suggest that the majority of other Pseudomonas and Acinetobacter spp. isolates clearly produced the carbapenem-hydrolyzing enzymes to gain their resistance to carbapenem, while P. aeruginosa isolates obtained other resistance mechanisms such as AmpC or extended spectrum β-lactamase and porin loss, as previously reported.13,14
Among the carbapenem-nonsusceptible isolates, MBL producers included only 6% (243 of 4077) of P. aeruginosa spp. and only 7% (262 of 3998) of Acinetobacter spp. Interestingly, 157 of 247 (64%) of the other Pseudomonas spp. produced MBL, and almost all of them (152 of 157) were P. putida, which was a much higher incidence than those of the other species. These findings suggest that P. putida can be a reservoir for MBL, as previously described.15 Acinetobacter spp. with different carbapenem-resistance mechanisms, such as OXA-type β-lactamases, have become prevalent.16
In P. aeruginosa, the incidence of MBL producers increased until the mid-2000s in this study, as shown in Japan,17 while in more recent years, these isolates gradually decreased in this study, as described in a previous report.18 Likewise, Cavalcanti, et al.19 reported that a higher prevalence of MBL-producing P. aeruginosa was observed in 2002-2003 in Brazil, while the level decreased significantly in 2008-2009, suggesting that the resistance to carbapenems by these recent P. aeruginosa isolates was not due to the spread of MBL-positive clones. In this study, P. aeruginosa isolates carrying a blaVIM-2-like gene were highly prevalent, comprising 90% to 100% of the P. aeruginosa strains in 2003 to 2004,5 although they were reduced to 34% while those with blaIMP-1-like genes increased to 69% in 2010.
Among MBL-producing Acinetobacter spp. isolates, the prevalence of blaIMP-1-like genes also increased to 85% in 2012. The range of strains carrying blaSIM-1-like genes remained low. Two isolates carrying blaNDM-1-like genes that were isolated in 2011 were identified as A. pitti and A. guillouiae. To our knowledge, this is the first report of a clinical isolate of A. guillouiae carrying the blaNDM-1-like gene. Most of the other Pseudomonas spp. isolates carrying MBL genes were identified as P. putida, and their MBL genes were blaVIM-2-like.
Among the 254 carbapenem-nonsusceptible Enterobacteriaceae, only five isolates, two K. pneumoniae, two E. cloacae, and one S. marcescens, produced MBL, suggesting that the major carbapenem resistance mechanism in Enterobacteriaceae was not MBL. Our results support previous reports that suggested that carbapenem resistance in Enterobacteriaceae was comediated with AmpC beta-lactamase and outer membrane protein loss in K. pneumoniae, E. cloacae, and S. marcescens.20,21,22
PFGE analysis revealed that the pulsotypes of IMP-6- and VIM-2-producing P. aeruginosa strains were clearly separated. The major pulsotypes in the IMP-6-producing P. aeruginosa were A2 and A3, while those in the VIM-2-producing P. aeruginosa were A1 and C1 types (Table 2). Likewise, the pulsotypes of Acinetobacter spp. isolates obviously differed according to MBL type (Table 2). These findings suggest that the plasmids carrying the MBL gene are not promiscuous, although they do have clone preference. Interestingly, Acinetobacter spp. isolates in the E1, H1, I4, J3, N1, and Q subgroups showed identical PFGE patterns, despite the differences in species. This suggests that the identification of Acinetobacter species is important for evaluating clonal outbreaks in hospital settings, as the misinterpretation of a clonal outbreak occurred among the different species.
Other mechanisms may block the cross-over of resistance plasmids between clones. Further study is warranted to elucidate this supposition. The hybridization of S1-digested DNA showed that the sizes of blaVIM-2 gene-carrying plasmids in P. aeruginosa, P. putida, and Acinetobacter spp. isolates were diverse (Supplementary Fig. 1, only online). It is noteworthy that the blaVIM-2 gene-carrying plasmids in Acinetobacter spp. were in multimer forms, indicating that the plasmids did not replicate themselves in the same way as with P. aeruginosa. Further plasmid sequence analysis using massive parallel sequencing technology has been undertaken.
In conclusion, MBL-producing clinical isolates of P. aeruginosa and Acinetobacter spp. were reduced, and carbapenemase-producing Enterobacteriaceae were found to be rare in Korea. Continuous surveillance studies and further deep sequencing are necessary to understand the dissemination mechanism of the carbapenem-nonsusceptible Gram-negative bacilli isolates in order to control their spread.

 XML Download
XML Download