

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
New neurons are generated from neural stem cells (NSC) throughout adulthood [1]. The generation of new neurons in adult brain is mostly associated with two regions: the subventricular zone (SVZ) of lateral ventricles and the subgranular zone (SGZ) of hippocampal dentate gyrus (HDG) [23]. NSCs in SVZ and SGZ are self-renewing. They are multipotent cells that give rise to neurons, astrocytes, and oligodendrocytes. They seem to have astrocyte like appearance. NSCs are known to express vimentin and glial fibrillary acid protein (GFAP) [45]. Neural progenitors and stem cells (NSCs) of the SVZ can differentiate into neuroblasts. They can migrate a long distance tangentially through rostral migratory stream and differentiate into interneurons in the olfactory bulb (OB). However, NSCs of the hippocampal SGZ can only migrate a short distance into granular cell layer where they differentiate into projection neurons [6].
Adult neurogenesis from HDG and SVZ can be divided into five stages: 1) proliferation, NSCs give rise to transiently amplifying cells and proliferating progenitor cells tightly associated with astrocytes and vascular structure; 2) differentiation and survival, transiently amplifying cells differentiate into immature neuron; 3) migration, immature neurons migrate a short distance into hippocampal granular cell layer and OB; 4) axon/dendrite targeting, immature neurons extend their projecting axon and short dendrites, and 5) synaptic integration, new granular neurons make synaptic integrations with other neurons [78]. Recent reports have shown that new neurons generated from neural stem cells might play pivotal roles in cognition and brain repair. For example, water maze exercise and enriched environment associated with improved memory function and synaptic plasticity can enhance adult neurogenesis in HDG [9]. In addition, adult neurogenesis is increased from HDG and SVZ after brain stroke, selective lesion, and seizure [1011]. However, roles of newborn neurons derived from HDG and SVG are currently unclear.
Toll-like receptors (TLRs), hallmarks of innate immunity, are increasingly implemented in central nervous system (CNS) plasticity-related processes including neurogenesis. They exhibit differential expression patterns in the brain with various functions in development. Critical roles of TLR3 [12] in NSCs and TLR4 [13] in microglia modulated proliferation and neural differentiation have been shown in adult brain [14]. TLR8 regulates neurotoxicity related to neural development [15] while TLR9 attenuates neurogenesis after seizure induced aberrant hippocampal neurogenesis [16]. Expression pattern of TLRs during brain development has also demonstrated roles of TLRs in neuronal plasticity and neurogenesis [17]. TLR2 is expressed in the central nervous system (CNS) [18] and peripheral nervous system (PNS) [19]. A recent study has shown that TLR2 is expressed abundantly in neurogenic niches of adult brain such as the hippocampus [20] of mammalian. It regulates adult hippocampal neurogenesis [21]. TLR2 can induce pro-inflammatory chemokines and pro-apoptotic proteins affected by microenvironment or damage [222324].
Transient whole brain ischemia can induce neuronal cell death in the brain [2526], notably in the hippocampal dentate gyrus. Adult neurogenesis in the dentate SGZ is affected by various physiological conditions, including CNS diseases, ischemia, epileptic, and seizure [2728]. Recent reports have shown that expression levels of TLR2 protein and mRNA in ischemic mouse brain are upregulated in TLR2 KO mice compared to those in WT mice following cerebral focal ischemia injury, while TLR4 KO mice exhibit reduced infarct size [2930]. Previously, we have shown that TLR4 plays a critical role in adult hippocampal neurogenesis of inflammatory-related neurological disorders [31]. However, the role of TLR2 in adult neurogenesis in global or focal cerebral ischemia is not well studied yet.
Thus, the objective of the present study was to elucidate the role of TLR2 in adult neurogenesis from neural stem cells of HDG in photochemical-induced cerebral ischemia.
METHODS
Animal preparation
All experiments were approved by the Animal Care and Use Committee of Chonnam National University. TLR2 knock-out (TLR2 KO) mice with C57BL/6 background mice were provided by Dr. Akira (University of Tokyo, Japan). Seven-week-old male C57BL/6 mice were purchased from Daehan Biolink (Chungbuk, Korea). Male C57BL/6 mice (body weight, 15–20 g) were caged in an air-conditioned room maintained at 22±2℃ with relative humidity of 50±10% and a 12/12 h light/dark cycle. Animals were provided free access to tap water and conventional mouse chow diet. They were acclimated for one week prior to the beginning of the study. Average age of mice used in this study was 7 weeks. Mice were divided into the following four groups: 1) Wild type, 2) TLR2 KO, 3) Wild type with photochemical-induced cerebral ischemia, and 4) TLR2 KO with photochemical-induced cerebral ischemia mice (n=5 mice/group).
Photothrombotic cerebral ischemia
Photothrombosis was performed using published procedure [32] with slight modifications. Briefly, Rose Bengal (Sigma, St. Louis, MO, USA) was dissolved in sterile saline at a concentration of 10 mg/ml. Then 0.1 ml of solution containing 1 mg of Rose Bengal was injected intravenously to each mouse five minutes before illumination. Mice were placed in a stereotactic frame. Their rectal temperature was monitored and controlled at 36.5–37.5℃ using a heating pad. At the dorsal aspect of the head, the skull was exposed by a median incision of the skin. The periosteum was gently removed to identify bregma and lambda points. Then 532 nm green laser light (1.5 mm aperture) was centered at 2 mm posterior and 2 mm laterally from the bregma. The duration of laser exposure was 10 minutes. Cerebral infarction was confirmed by 2,3,5-triphenyltetrazolium chloride staining on the next day.
BrdU injection
Bromodeoxyuridine (BrdU; Sigma, St. Louis, MO, USA) was dissolved in physiologic saline containing 0.9% NaCl and injected intraperitoneally into mice (50 mg/kg). To examine proliferation of neural stem cells in hippocampal DG, BrdU was injected once daily into TLR2 KO mice and WT mice. Mice were then sacrificed at three hours after BrdU injection. To study the survival of newborn cells from neural stem cells and differentiation rate of neural stem cells, BrdU was injected once daily for five consecutive days. Mice were then sacrificed at 28 days after the final BrdU injection.
Tissue preparation
Animals were perfused transcardially with 0.1 M phosphate-buffered saline (PBS, pH 7.4) and fixed with fresh and cold 4% paraformaldehyde (PFA) in 0.1 M PBS. Brains were dissected and post-fixed overnight with 4% PFA in 0.1 M PBS at 4℃, washed for 6 h in PBS at 4℃, and cryoprotected overnight with 30% sucrose in 0.1 M PBS at 4℃. Brains were then embedded in freezing media (optimum cutting temperature), frozen in chilled isopentane (−25℃), and stored at −80℃ until sectioning. Brains were cryocut sagittally or coronally to sections of 40 µm in thickness using a cryostat (model CM3050 S; Leica Microsystems, Bannockburn). Sections were stored in cryoprotectant solution (25% ethylene glycol, 25% glycerol, and 0.05 M sodium phosphate buffer, Na-PB) at −20℃.
Immunohistochemical fluorescent staining
Sections were washed with Na-PB and mounted onto charged slides. Sections stained for BrdU were pretreated with 2 N HCl at 37℃ for 30 min and neutralized with PBS before incubation with primary antibody. Sections were blocked with PBS containing 5% horse serum and 0.4% Triton X-100 in PBS (PBST) for 60 min followed by incubation with primary antibody in the same buffer solution at 10℃ overnight. Primary antibodies were used at the following concentrations: rat anti-BrdU (1:200; Abcam, Cambridge, UK), mouse anti-GFAP (1:200; Chemicon, Temecula, CA, USA), mice anti-neuronal nuclear antigen (NeuN, 1:100; Chemicon), rabbit anti-ionized calcium-binding adapter molecule 1 (Iba-1, 1:100; Wako Chemicals USA, Inc., Richmond, VA, USA), and goat anti-Doublecortin (DCX, 1:100; Cell signaling, Danvers, MA, USA). Sections were washed three times with PBST for 10 min at room temperature and blocked in PBST containing 5% horse serum for 30 min. Samples were then incubated with secondary antibodies conjugated to FITC (green, Jackson Immuno-Research, West Grove, PA, USA) or CY3 (red, Jackson Immuno-Research) at room temperature for two h, washed three times with PBST, stained with 10 mg/ml 4′, 6′-diamidino-2-phenylindole (DAPI) (Sigma) for 10 min, and coverslipped.
Cell death analysis
To examine apoptosis of NSCs in the hippocampal SGZ, immunofluorescent staining was performed using activated caspase-3 antibody (1:200, Cell Signaling, Danvers, MA, USA) according to the immunofluorescent staining method described above. Activated caspase-3 positive cells were counted throughout the hippocampal SGZ of six serial sections.
Quantification and image analysis
Staining was analyzed with a confocal microscope (Zeiss, Thornwood, NY, USA) equipped with an argon/krypton laser (488 nm), two helium/neon lasers (543 and 633 nm), a Coherent (Santa Clara, CA, USA), and a Chameleon two-photon laser at 725 nm to image DAPI staining. Pictures were obtained with a Spot RT Slider camera and associated software (version 3.4; Diagnostic Instruments, Sterling Heights, MI, USA) or LSM software (Zeiss, Thornwood, NY, USA). Co-localizations were investigated by performing z-stack acquisitions and three-dimensional reconstructions. Images were analyzed with Image J (Ver.1.4, NIH, USA). Adobe Photoshop version 7.0.1 (Adobe Systems, San Jose, CA, USA) was used to adjust contrast and brightness.
Data analysis
A complete series of one in ten sections of the DG was analyzed. For each experiment, nine sections per mouse were selected for analysis. Immunolabeled cells were counted and multiplied by 10 to obtain the total number of labeled cells throughout the DG. Data are expressed as mean±SEM. Statistical analyses were performed using one-way analysis of variance (ANOVA) followed by Student-Newman-Keuls test for multiple comparisons. Results were considered statistically significant when p value was less than 0.05 [33].
RESULTS
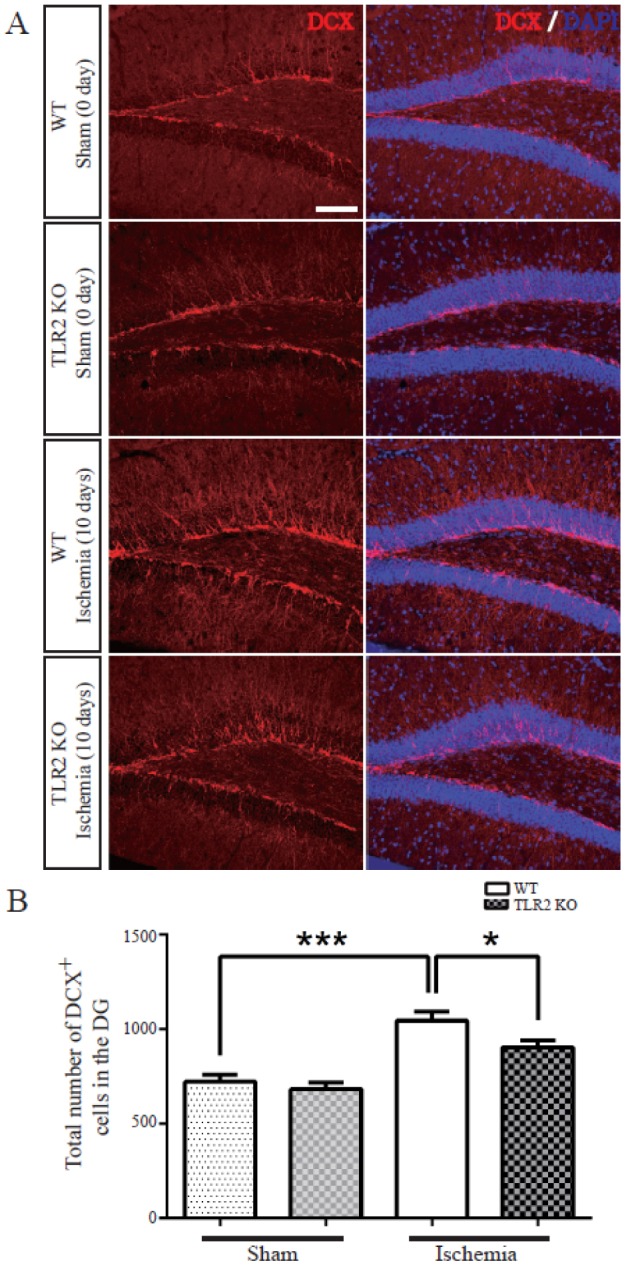
TLR2 promotes neuronal differentiation of neural stem cell in the dentate gyrus after photothrombotic ischemic injury
To investigate whether TLR2 could regulate neurogenesis in the dentate gyrus after photothrombotic ischemia, WT and TLR2 KO mice were sacrificed at 10 days after ischemia and their brain sections were stained with DCX, an immature new born neural marker. The number of DCX positive cells was not significantly different between WT and TLR2 KO mice under normal condition. After ischemic injury, DCX positive cells were increased in WT (32.4±5.9% increase) and TLR2 (22.4±4.9% increase) groups compared to those under sham condition. However, DCX positive cells were significantly reduced in TLR2 KO mice compared to those in WT mice after ischemic injury (14±5.7% decrease, p=0.0387) (Figs. 1A and B), indicating that DCX-positive cells in SGZ were significantly increased by photothrombotic ischemia in both WT mice and TLR2 KO mice. However, the number of DCX-positive cells in TLR2 KO mice was decreased compared to that in WT mice after ischemia.
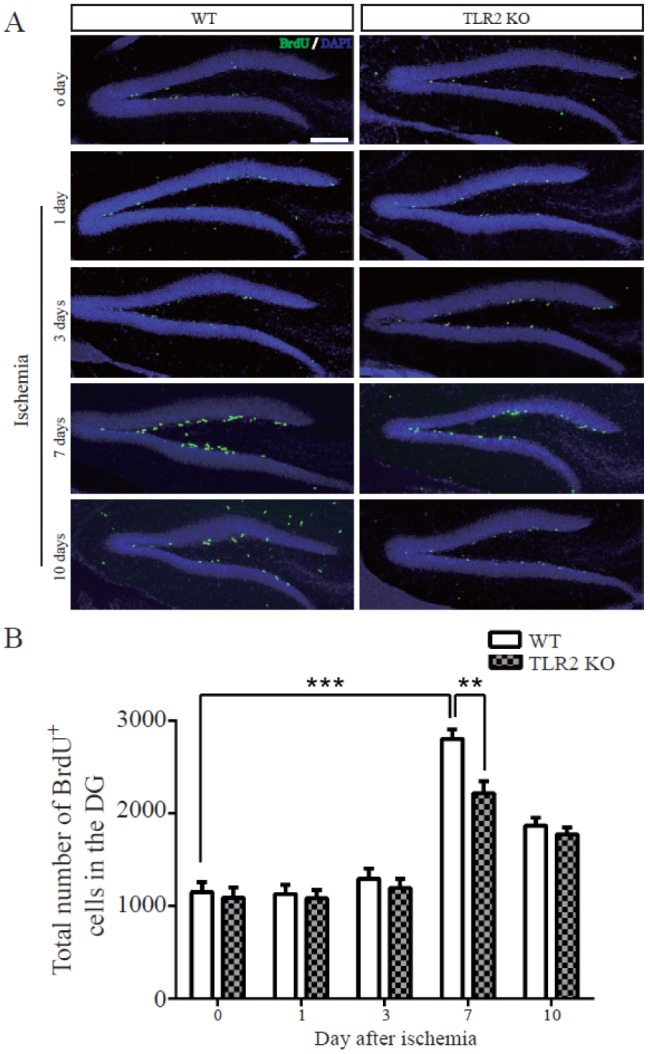
TLR2 increases cell proliferation in adult dentate gyrus after photothrombotic ischemia
BrdU-labeled nuclei in SGZ of both control and ischemic brains appeared as irregularly shaped clusters located exclusively in the SGZ. This pattern is a characteristic of dentate gyrus progenitor cells. The number of BrdU-positive cells was not changed by sham operation or at 1 or 3 days after photothrombotic ischemia (Fig. 2A), demonstrating that the stress of surgery did not increase cell proliferation. The number of BrdU-positive cells was gradually increased in both TLR2 KO mice (113.7±6.9% compared to WT at 0 day) and WT (167.7±6.4% compared to TLR2 KO at 0 day) mice at 7 days after photothrombotic ischemia, with 2.5-fold increase in BrdU immunoreactive nuclei per square millimeter dentate gyrus compared to sham operating group (Fig. 2B). The increasing rate of BrdU-positive cells in TLR2 KO mice (58.5±7.3%) was decreased compared to that in WT mice. These results demonstrated that TLR2 stimulated the proliferation of adult NSCs after photothrombotic ischemia in the hippocampal DG in vivo.
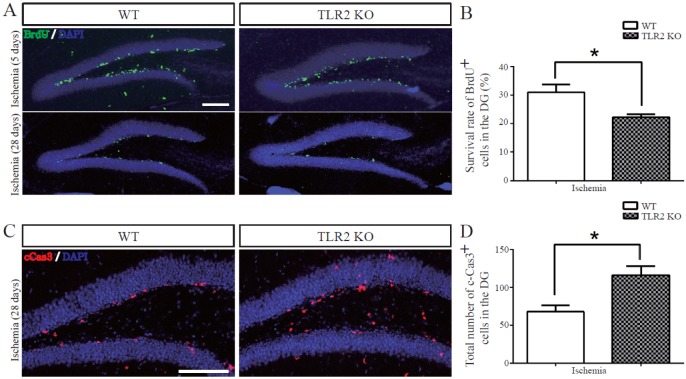
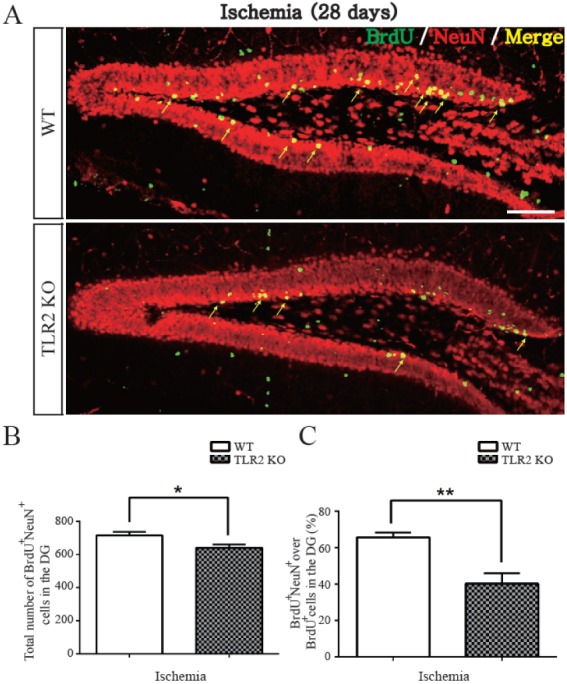
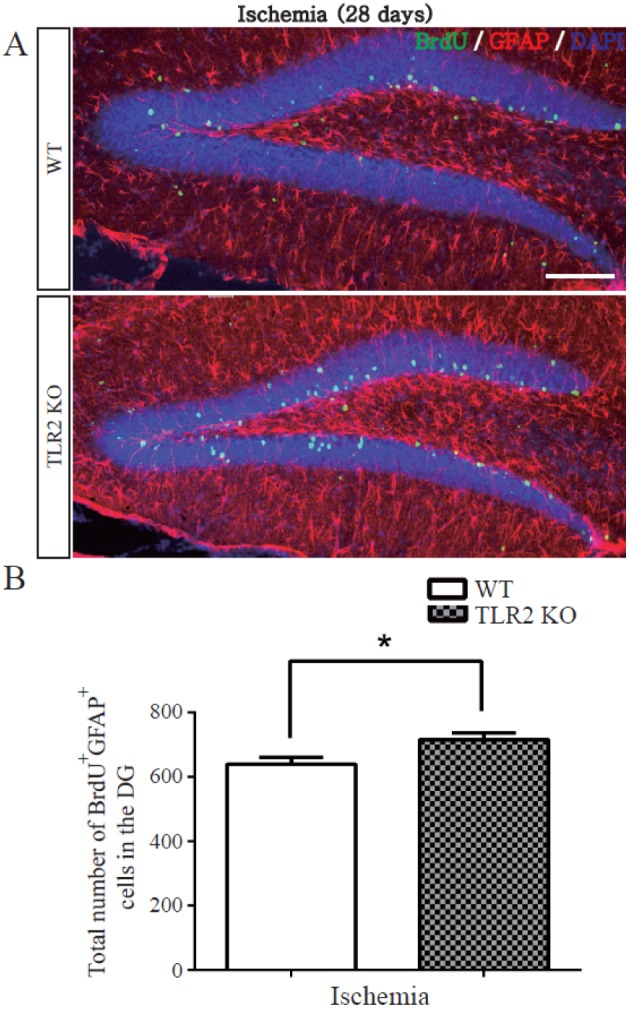
TLR2 promotes survival of newborn cells and neuronal differentiation from neural stem cells after photothrombotic ischemia
To examine the survival and matured neuronal differentiation of BrdU-positive cells after ischemic injury, numbers of BrdU-positive cells and NeuN/BrdU or GFAP/BrdU double-positive cells were determined at 28 days after BrdU injection once daily for five consecutive days. NeuN was used as a marker of mature neuron while GFAP was used as a marker of glial cell. BrdU positive cells at 28 days after five days of BrdU injection were diminished in the hippocampal DG of TLR2 KO mice (8.8±3.0% decrease, p=0.0182) compared to those in WT mice (Figs. 3A and B). To investigate apoptosis of neural stem cells or newborn neuron in the hippocampal DG, activated caspase-3 (an apoptotic executing caspase) immunostaining was performed. Caspase 3-positive cells in SGZ were increased in TLR2 KO mice (9.6±2.9% increase, p=0.0165) compared to those in WT mice (Figs. 3C and D), indicating that new born neurons underwent apoptosis. The number of NeuN/BrdU double-positive cells in the hippocampal DG of TLR2 KO mice were less than those in WT mice (7.6±3.0% less compared to WT, p=0.0371, Fig. 4B). In addition, the percentage (%) of NeuN/BrdU double-positive cells over the number of BrdU positive cells in the hippocampal DG of TLR2 KO mice were less than those in WT mice (25.5±6.3% less compared to WT, p=0.0067, Fig. 4C). However, the number and percentage of GFAP/BrdU positive cells in TLR2 KO mice were higher (7.6±3.0% higher) than those in WT mice (Figs. 5A and B). These results indicated that TLR2 enhanced neuronal differentiation but inhibited astrocyte differentiation from stem cells of hippocampal dentate gyrus after ischemic injury, suggesting that TLR2 could modulate the fate of neural or glial differentiation from neural stem cells after ischemic injury.
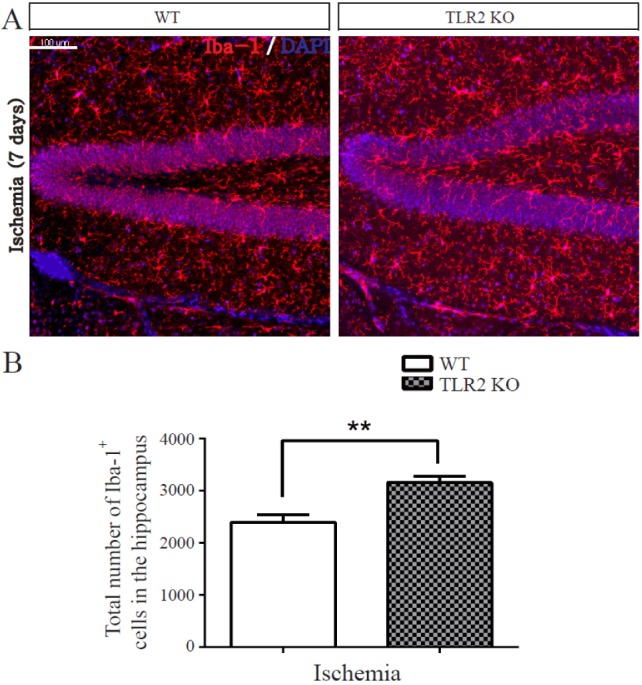
TLR2 inhibits microglia activation after photothrombotic ischemia
Recent brain studies have shown that microglia express TLR2 and modulate hippocampal neurogenesis through secreting immune related cytokines and neurotrophic factors [34]. To determine the effect of TLR2 on ischemia-induced inflammation, the activity of microglia following ischemia was quantified by measuring immunoreactivity of Iba-1, a specific marker of microglia (n=5 each). The number of Iba-1-positive cells in DG was increased in TLR2 KO mice compared to that in WT mice at 7 days after photothrombotic ischemia (76.23±18.8% increase, p=0.0066, Fig. 6A and 6B). This result demonstrated that TLR2 could suppress the activity of microglia in the hippocampus after ischemia.
DISCUSSION
Toll was first discovered in Drosophila melanogaster. It regulates dorso-ventral patterning during embryonic development and controls immune responses [3536]. After the discovery of TLR4, a mammalian homolog of Toll, many mammalian homologs have been identified across diverse species. In mammalian, TLRs are involved in innate immune activity and activation of the adaptive immune system. They also have physiological and metabolic roles in adults. There was a report that Knock-out of TLR2 affects immune response through modulating proinflammatory cytokines in human coronary artery endothelial cells [37]. Human adipose-derived stem cells (hASCs) expressing all TLRs including TLR2 are activated by TLR ligands such as LPS, leading to significant upregulation of IL6 and IL8 that can stimulate osteoblast differentiation [38]. Mesenchymal stem cells derived from human turbinate tissue also express TLR2 and TLR4 while TLR agonist affects proliferation, differentiation, and immune responses [39]. TLR2 regulates multiple metabolism along with TLR4 to control autonomic nervous system [40].
Adult neurogenesis continues in the hippocampal DG and SVZ throughout adulthood for cognition or repair of lost neurons following injury or disease [4142]. Endogenous neurogenesis can be regulated by various stimuli, including physical exercise, enriched environment, stroke, and growth factors [4344]. Recent reports have shown that ischemic brain injury upregulates vascular endothelial growth factor (VEGF), fibroblast growth factor (FGF)-2, and glial derived neurotrophic factor (GDNF) expression, resulting in increased neurogenesis from adult dentate gyrus [454647]. In the present study, photochemical-induced cerebral ischemia also enhanced neurogenesis in hippocampal dentate gyrus in both WT mice and TLR2 KO mice. This suggests that cerebral ischemia is one of the major stimuli for neurogenesis from neural stem cells. TLR2 is known as a candidate receptor that may regulate adult neurogenesis from neural stem cells in SVZ and hippocampal SGZ [4849]. However, whether TLR2 regulates neurogenesis under cerebral ischemia condition in hippocampus has not been reported yet.
In general, adult neurogenesis can be divided into five separated stages: stem cells proliferation, survival, migration, differentiation, and synaptic integration. In the present study, the number of DCX (newborn neuronal marker)-positive cells was decreased in hippocampal DG TLR2 KO mice compared to that in WT mice after cerebral ischemia, indicating that TLR2 could promote neurogenesis under cerebral ischemic injury. To elucidate the mechanism underlying the stimulatory effect of TLR2 on adult neurogenesis from neural stem cells after cerebral ischemic injury, stem cells proliferation, cell survival, and differentiation were examined in the present study. The number of BrdU-positive cells was lower in hippocampal DG of TLR2 KO mice than that in WT mice after cerebral ischemic injury, demonstrating that TLR2 could promote proliferation of neural stem cell in hippocampal DG, resulting in increased neurogenesis in hippocampal DG after cerebral ischemia. Our analysis using TLR2 KO mice after brain specific damage demonstrated that TLR2 played an important role in brain development and self-renewal property of hippocampal neural stem cells. It has been recently reported that TLR2 activation upregulates cell cycle related molecular targets through modulating inflammatory mediators in stem cells [50]. However, such data are not consistent with a previous study showing that TLR2 is not involved in the regulation of neural stem cell proliferation in hippocampal DG without cerebral ischemia [48]. In addition, the rate of neural differentiation from neural stem cells was compared between hippocampal DG of TLR2 KO mice and WT mice after cerebral ischemia in the present study. The percentage of NeuN (mature neural marker)/BrdU-positive cells were found to be lower in hippocampal DG of TLR2 KO mice than that in WT mice after cerebral ischemia, whereas the percentage of GFAP (astrocyte marker)/BrdU-positive cell was higher in hippocampal DG of TLR2 KO mice compared to that in WT mice after cerebral ischemia. These results suggest that TLR2 can enhance neural differentiation from neural stem cells in DG after cerebral ischemia, resulting in increased neurogenesis after cerebral ischemia. This is consistent with results of a previous report showing that TLR2 increases neuronal differentiation in mouse embryonic neural stem cells [48].
To determine whether TLR2 receptor was associated with apoptosis or survival of newborn neuron, the number of activated caspase-3-positive cells was examined. In the present study, the number of BrdU-positive cells at 28 days after consecutive BrdU injection for five days was lower in the hippocampal DG of TLR2 KO mice compared to that in WT mice after cerebral ischemia. Besides, the number of caspase 3-positive cells was higher in the hippocampal DG of TLR2 KO mice compared to that in WT mice after cerebral ischemia. This result supports that TLR2 has a protective effect on the survival of adult NSCs at a late step which is reduced by ischemia. It is consistent with a previous report demonstrating that TLR-mediated NF-κB activation seems to have anti-apoptotic role in neuron under ischemic injury or oxidative stress condition [515253]. However, TLR2 can regulate anti-apoptotic or cytotoxic function in various cell types through NF-κB signaling pathway [5455]. In the present study, the recruitment of activated microglia was evaluated under ischemic condition. Activated microglia was found to be significantly increased in TLR2 KO mice compared to that in WT mice, meaning that TLR2 decreased microglia activation. Taken together, our findings suggest that TLR2 can augment cell survival of newborn neural cells in the hippocampal DG through modulating microglia activity after cerebral ischemia.
TLR2 is known to be expressed in all tissues, including adult neural stem cells, microglia, and neurons [565758]. However, the present study did not study tissue specific effect of TLR2 because whole body knock-out mice were used in this study. It has been reported that Cre recombinase under control of seleted tissue promoter such as nestin or Sox2 promoter can allow knock-out or overexpression of a floxed target gene [59]. Therefore, further study is needed to determine tissue specific effects of TLR2 through knockout or overexpression of TLR2 using Cre system in neural stem cells of tissue specific transgenic mice. Specific system such as inactivation of TLR2 in specific tissues by Cre recombinase might be used to regulate proliferation, differentiation, and survival of neural stem cells in hippocampal neurogenesis.
In summary, TLR2 receptor can enhance adult neurogenesis from neural stem cells in the hippocampal DG after cerebral ischemia through promoting proliferation and neural differentiation of neural stem cells and survival of newborn neurons. Therefore, TLR2 pathway that regulates hippocampal neurogenesis after ischemia might be targeted as a prospective therapeutic strategy to treat brain nerve injury and various neurodegenerative disorders.
 XML Download
XML Download