

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Parkinson's disease (PD) is known as a progressive neurological incurable disorder with no preventative nor effective long-term treatment strategies [1]. The phenotypes of PD are characterized by the progressive loss of dopaminergic neurons in the substantia nigra (SN); dopamine depletion in the striatum; and motoric instabilities including bradykinesia, rigidity, and tremor [2]. In most cases, the etiology of PD is not completely understood. However, several genes have been related to some cases of PD [3]. The presynaptic protein, α-synuclein (α-syn) gene, is a major component of intracytoplasmic inclusions termed Lewy bodies which are the pathological hallmarks of PD, and missense mutations in α-syn gene such as A53T, A30P, and E46K were found to be linked to a familial mode of PD [456]. Overexpression of WT or mutant A53T human α-syn in mice resulted in neuronal degeneration and motor deficits [7]. Although human α-syn accumulates within cell bodies of SN, locus cereleus (LC) and the olfactory bulb (OB), the mutant human α-syn (A53T) mice do not exhibit overt neuropathology. Moreover, the number of nigral neurons and the levels of striatal dopamine (DA) were unchanged relative to WT littermates up to 1 year of age [8]. Distinguished from the previous α-syn A53T mouse model, our lab has generated new PD mouse models; hemi and homo α-syn A53T mice exhibiting medium and high dopaminergic neuron loss, using 5 days of daily sub-toxic (10 mg/kg) injection of 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP). The levels of tyrosine hydroxylase (TH), synuclein, synaptophysin, DJ-1 and Bcl-2 expression levels in the nigrostriatal regions and behavioral tests in the hemi or homo transgenic mice expressing mutant A53T α-syn and their WT littermates were also assessed following MPTP treatment. With the help of MPTP-treated mutant A53T α-syn expressing Tg and WT mice, our study unravels the possible mechanism through which α-syn accumulation leads to Parkinson's disease.
METHODS
Transgenic mice
Protocols for animal use were reviewed and approved by the Institutional Animal Care and Use Committee at the Dongguk University Ilsan Hospital (No. 2013-1193) and were in accordance with National Institute of Health guidelines. Hemizygous (Prnp-SNCA*A53T) 83Vle mice with a B6C3H background were obtained from the Jackson Laboratory (Bar Harbor, Maine, USA) and were bred at the Dongguk University. From the offspring, wild-type, hemizygous and homozygous male mice were used for the experiments. Identification of the (Prnp-SNCA*A53T) 83Vle transgenic and non-transgenic mice was performed according to the results of the PCR with primers specific for the transgene sequence obtained from Bioneer (Daejeon, Korea), and further identification of zygosity was analyzed using hemizygous and homozygous DNA samples from F0 mice. Animals were maintained under constant temperature (22±1℃), relative humidity (55±1%), and 12 h light/12 h dark cycle (light on at 7:00 am). They were given tap water and commercial rodent chow (Samyang Feed, Daejeon, Korea) ad libitum. Wild-type mice (n=21), A53T hemizygous mice (n=21) and A53T homozygous mice (n=21) at 4~5 months of age were assigned to three groups: control (saline treatment), sub-toxic dose of MPTP (10 mg/kg) and toxic dose MPTP (30 mg/kg) at 7 mice in each group. Saline or MPTP was daily treated through intraperitoneal (i.p.) injection for 5 days, and behavior tests were performed at 12 day. Animals were decapitated after behavior test.
DNA isolation and genotyping
DNA samples were extracted from the ears of test mice using a Phenol-Chloroform Isoamyl Alcohol (PCI) DNA extraction method [9]. For validation purposes, extracted DNA samples were genotyped according to the standard PCR protocols obtained from the Jackson Laboratory. For further investigation of zygosity, Real Time PCR was performed using Taqman qPCR protocols obtained from the Jackson Laboratory.
Immunoblotting
The SN tissues were removed and homogenized in lysis buffer containing 50 mM Tris-base (pH 7.5), 150 mM NaCl, 2 mM EDTA, 1% glycerol, 10 mM NaF, 10 mM Na-pyrophosphate, 1% NP-40, and protease inhibitors (0.1 mM phenylmethylsulfonylfluoride, 5 µg/ml aprotinin, and 5 µg/ml leupeptin). Lentivirus infected SH-SY5Y cells were harvested in lysis buffer. The tissue or cell lysates (30 µg) were electrophoresed using sodium dodecyl sulphate-polyacrylamide gels and transferred to nitrocellulose membranes and then incubated with anti-TH (Santa cruz Scientific,USA, 1:1000, Rabbit), anti-DJ-1 (Merck Millipore, Carolina, USA, 1:1000, Rabbit), anti-Bcl-2 (Cell signaling technology, Beverly, MA, USA, 1:1000, Rabbit), anti-Synaptophysin (Abcam, Cambridge, UK, 1:1000, Rabbit), anti-α-synuclein (GeneTex, Irvine, CA, USA, 1:1000, Rabbit), anti-UBE2N and anti-β-actin (Cell signaling technology, Beverly, MA, USA, 1:1000, Rabbit) for 16 h at 4℃. After washing with Tris-buffered saline-Tween 20 (TBST) (0.05%), the blots were incubated with horseradish peroxidaseconjugated anti-rabbit (1:5000) or anti-mouse (1:5000) IgG (Thermo Fisher Scientific, USA), and the bands were visualized using the ECL system (Thermo Fisher Scientific, USA). Band images were obtained by using a Molecular Imager ChemiDoc XRS+ (Bio-Rad, Hercules, CA, USA), and band intensity was analyzed using Image Lab™ software version 2.0.1 (Bio-Rad, Hercules, CA, USA).
Immunostaining
The mice were perfused transcardially with phosphatebuffered saline (PBS) followed by 4% paraformaldehyde in 0.2 M phosphate buffer. The brains were removed, post-fixed, and cryoprotected. For immunohistochemistry, brains (40 µm thick) were stained with anti-TH antibody (Santa Cruz Scientific, USA) followed by Alexa Fluor 488 secondary antibody (Thermo Fisher Scientific). Biotechnology; sc-253), followed by Alexa Fluor 488 (1:200; Thermo Fisher Scientific; A-21206). The sections were observed under an Olympus FV-1000 laser confocal scanning.
Adhesive removal test
The adhesive removal test was established in α-syn A53T mice to determine the effects of MPTP treatment on sensory-motor behavior. During a test, cage mates were placed in a clean cage while the test mouse was placed in its home cage. For testing, a small adhesive patch (circular 0.3 cm diameter) was applied to each snout using a pair of small forceps, and the time to contact and remove each adhesive tape was recorded. The latency to contact the patch with the paw was used to assess sensory impairment. If the mouse did not contact or remove the sticker within 60 s then the trial was ended and the sticker was removed manually by the experimenter. All mice received 3 trials.
Beam traversal test
Motor coordination and balance in 4~5 months of α-syn A53T and WT littermate mice was tested using a challenging beam traversal test. In brief, a plastic beam, 1 m length total, and comprising of four (25 cm length) sections that gradually decreased in diameter from 3.5 cm to 0.5 cm in 1 cm increments was used. Animals were trained to traverse the beam (from widest to narrowest) directly into the animal's home cage. Each mouse received two days of training (5 trials each) followed by testing on the third day. During the testing phase, a wire mesh grid (1 cm2) of corresponding beam width was placed over the beam. Animals were videotaped while traversing the beam, over 5 trials. Video records were analyzed at slow motion, by an investigator blinded to the genotype of the animals and the time taken to traverse the beam, and number of foot slips off the beam were determined over the 5 trials and averaged.
Cell culture works
SH-SY5Y cells were obtained from the American Type Culture Collection (Rockville, MD, USA) and cultured in DMEM containing 10% of FBS and 1% of antibiotics (Hyclone Laboratories Inc., Logan, UT, USA). To investigate the effect of α-syn A53T expression in cell viability, cells were infected with 1, 5, or 10 m.o.i of α-syn A53T lentivirus for 3 days and incubated with 1 mg/ml of 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) (Sigma) at 37℃ in a CO2 incubator for 3 h. The formazan dye was eluted using DMSO. The plate was shaken and measured using a spectrophotometer (iMark™ Microplate Absorbance Reader, Bio-Rad, Hercules, USA) at a wavelength of 570 nm.
Virus production
Human cDNA of α-syn was obtained by PCR of cDNA of human brain and α-syn A53T was generated by site-directed mutation system (Thermofisher Scientific, USA) and pCDH-α-syn A53T lentivector construct was generated by ligating α-syn A53T PCR product into a lentiviral backbone pCDH (System Bioscience, Palo Alto, CA, USA). The construct was sequenced and ensure that no PCR-based mutations were generated. Lentivirus of α-syn A53T were produced and concentrated by manufactural manual (System Bioscience, Palo Alto, CA, USA). The virus titer was determined by infecting 293TN cells with serial dilutions of virus and counting the number of GFP or α-synuclein positive cells (as assessed by immunocytochemistry). A multiplicity of infection (m.o.i.) was calculated and 1, 5, or 10 m.o.i was used to infect SH-SY5Ycells.
RNA interference
RNA oligonucleotides for the knockdown of human DJ-1 (Cat. 4390824) (DJ-1 siRNA) and scrambled siRNA (Control siRNA) were purchased from Invitrogen Stealth siRNA by using BLOCKiT™ RNAi designer (Carlsbad, CA). Cells were plated onto 12-well plates (5×105 cells/well) and allowed to adhere for 24 h before siRNA transfection using Effectene transfection reagent (Qiagen, CA), according to the manufacturer's instructions. At 48 h after transfection of siRNA, cell viability was measured by MTT assay and the cells were harvested for western blotting.
Statistical analysis
All statistical analyses were accompanied with SPSS (ver. 21, Somers, NY, USA). All the amount were expressed as means±SD. The data were analyzed by one-way ANOVA, and the difference among means were analyzed using the Dunnett's test or Tukey– Kramer's multiple comparison test. Differences were considered significant at p<0.05.
RESULTS
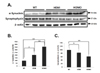
Evaluation of synaptophysin and synuclein expression in the SN of WT and α-syn Tg mice
PD is linked to abnormal aggregation of α-syn [7101112] and α-syn is known to play a role in synaptic transmission [13]. As shown in Fig. 1, α-syn expression was increased in α-syn Tg mice compared to that of WT. The α-syn expression level was correlated with hemi and homo α-syn Tg (Fig. 1B).
Furthermore, a recent study demonstrated that increased expression of mutant A53T α-syn displays decreased synaptophysin, a major synaptic vesicle protein related with synaptic connection [14]. Accordingly, the level of synaptophysin was decreased dependently to the level of α-syn (Fig. 1A, C). These results indicate that overexpression of α-syn may interrupt the synaptic transmission by down-regulating synaptophysin.
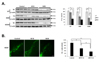
Evaluation of TH expression in the SN of MPTP-treated Tg mice
Overexpression of mutant α-syn has been reported to increase the susceptibility of neuronal cells to oxidative injury [1516]. Toxic agent such as MPTP or 6-OHDA is used as an oxidative injury. In order to develop the Parkinson's disease animal model in α-syn Tg mice showing impaired motor function as well as dopamine cell loss in SN, sub-toxic dose (10 mg/kg) or toxic dose (30 mg/kg) of MPTP was treated in WT, hemi and homo α-syn Tg mice. As an indicator of SN DA level, tyrosine hydroxylase (TH) protein levels in the SN of WT or Tg mice were observed via Western blot. Following MPTP treatment, TH expression levels were significantly decreased in toxic dose but not in subtoxic dose in WT mice (p<0.05, Fig. 2A). However, when α-syn Tg mice were treated with MPTP, TH expression in both hemi and homo α-syn Tg mice exhibited a dose-dependent reduction and in homo α-syn Tg mice, 30 mg/kg MPTP injection showed the highest reduction level of TH in SN. Our findings suggest that TH levels are reduced in SN of α-syn Tg mice but not in WT mice by sub-toxic dose (10 mg/kg) of MPTP injection, and homo A53T α-syn Tg mice showed increased vulnerability towards MPTP injection compared to that of hemi α-syn Tg mice. To confirm the TH-positive cells in the SN of hemi A53T α-syn Tg mice, immunostaining of TH was used. As shown in Fig. 2B, the number of TH positive cells was decreased in a dose-dependent manner. Moreover, we examined the expression level of Bcl-2, an antiapoptotic protein in the SN. The levels of Bcl-2 were normalized to β-actin to compare the expression in hemi and homo α-syn Tg mice, and the graph showed that homo α-syn Tg mice exhibited significantly decreased Bcl-2 level than hemi α-syn Tg mice (Fig. 3). These results suggest that sub-toxic (10 mg/kg) MPTP injection is enough to induce oxidative injury causing cell death.
Behavioral tests of MPTP-treated WT or Tg mice
To evaluate the relationship of dopamine cell death and behavioral impairment, adhesive removal and challenging beam tests were used in MPTP-treated WT or Tg mice. In the adhesive removal test (Fig. 4A), dose-dependent treatment of MPTP in WT, hemi and homo A53T α-syn mice showed a trend towards longer latency but it was not significantly different. However, homo α-syn Tg mice showed significantly longer latency compared with 30 mg/kg of MPTP.
In the challenging beam test (Fig. 4B), control hemi and homo α-syn Tg groups showed a trend towards increased latency compared to that of WT control group; but not significantly different. When treated with 10 mg/kg of MPTP, hemi and homo α-syn Tg mice exhibited significantly longer latency compared to that of WT control. Furthermore, 30 mg/kg of MPTP treatment in all WT, hemi and homo groups induced significantly longer latency.
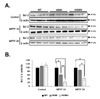
Evaluation of DJ-1 and UBE2N expression in the SN of WT or Tg mice
In order to find out responsible proteins related with vulnerability of mutant α-syn expressed neurons, DJ-1 and ubiquitin enzyme expression were examined. DJ-1, a ubiquitously expressed protein oxidative stress response protein [171819], is known to reduce the accumulation and toxicity of α-syn [181920]. The expression level of DJ-1 was significantly decreased in both homo and hemi α-syn Tg mice compared to that of WT (Fig. 5). Homo α-syn Tg mice exhibited slightly more decreased expression of DJ-1 than that of hemi α-syn Tg mice. These results demonstrate that the mutant A53T α-syn expression is associated with a decrease of DJ-1 level.
Next, we examined UBE2N (ubiquitin conjugating enzyme) expression level in the SN of mutant α-syn Tg mice. UBE2N expression was significantly decreased in hemi or homo α-syn Tg mice (Fig. 5). These suggest that overexpression of mutant α-syn may reduce the activity of these enzymes by down-regulation.
Evaluation of mutant A53T α-syn expression and DJ-1 down-regulation in the SH-SY5Y cells
To investigate the relationship of mutant A53T α-syn expression and DJ-1 down-regulation, SH-SY5Y cells were treated with 10 m.o.i. of mutant A53T α-syn lentivirus for 72 h. In the cells, α-syn expression was increased by two fold (Fig. 6A, upper panel). Then, the cells were treated with 0, 25, 50 or 100 µM of 6-OHDA for 24 h and the cell viability was examined by MTT methods. A53T α-syn overexpression significantly induced cell death by 20.03±9.9% (p<0.05) and when 100 µM of 6-OHDA was treated into the cells, cell viability was decreased by 49.74±8.3% (Fig. 6A). Next, we examined DJ-1 expression in the A53T α-syn expressed cells. As shown in Fig. 6B, as the expression of A53T α-syn increased, the expression of DJ-1 decreased, suggesting that A53T α-syn may down-regulate DJ-1 expression responsible for the cell survival. To confirm the contribution of DJ-1 repression in the cell viability, siRNA of DJ-1 was used. DJ-1 knock down completely induced cell death without toxin insult in the SH-SY5Y cells (Fig. 6C). Thus, these data suggest that the vulnerability or toxicity to oxidative injury such as MPTP or 6-OHDA of A53T α-syn overexpression can be explained by DJ-1 downregulation.
DISCUSSION
A53T α-syn Tg mice are known to express extensive mutated α-syn expression throughout the brain, including the SN, hippocampus, and neocortex [21]. A53T α-syn mutation exhibits aggregation of α-syn and is more toxic to neurons than WT α-syn [22]. α-syn is especially localized in the presynaptic nerve terminals [23]. Scott et al. reported that the higher-expressing α-syn displayed synaptic accumulation of α-syn accompanied by decreased synaptophysin [14]. In the same context, we confirmed that the increased A53T α-syn expression reduced synaptophysin level in a dose-dependent manner. These indicate that increased α-syn accumulation shows neuronal toxicity to the nearby protein activity.
Some studies have also implicated that the expression of A53T and A30P mutant α-syn in neuronal cell lines enhanced the susceptibility of these cells to oxidative insults such as H2O2 or 1-methyl-4-phenylpyridinium (MPP+), the active toxic metabolite of MPTP [15242526272829]. In vivo experiment reported that daily MPTP-treatment (30 mg/kg, i.p.) for 5 days in both mutant A30P α-syn expressing Tg and WT mice groups induced significant striatal DA depletion [30]. In our study, we used identical schedule and regimen in mutant A53T α-syn expressing Tg and WT mice and produced a similar result; toxic dose (30 mg/kg) of MPTP treatment induced severe SN TH expression depletion in Tg mice.
Thus, we attempted a sub-toxic dosage approach that was less damaging. Richfield et al. indicated that expression of human α-syn resulted in increased sensitivity to mild MPTP-treatment in Tg mice when using a low dose (15 mg/kg) of MPTP [12]. Yu et al. also revealed that sub-lethal administration of MPTP (15 mg/kg) for 5 days increased sensitivity of human α-syn expressing Tg mice that was demonstrated by a greater decrease in locomotor activity [31]. In this study, we injected 10 mg/kg of MPTP intraperitoneally for 5 days in a row in mutant A53T α-syn expressing Tg and WT mice, and our data showed that 10 mg/kg of MPTP treatment induced dopaminergic toxicity in hemi and homo α-syn Tg mice but not in WT. In addition, homo α-syn Tg mice exhibited higher sensitivity to MPTP administration.
Moreover, our behavior tests provided a supportive evidence that mutant A53T α-syn increases sensitivity to MPTP. In the adhesive removal test, time to remove paper was significantly increased in homo α-syn Tg mice, confirming that α-syn accumulation enhances vulnerability to MPTP. In the challenging beam test, hemi and homo α-syn Tg mice spent significantly longer time to traverse when treated with 10 mg/kg of MPTP as compared to that of WT. Cumulatively, our findings may suggest a new PD animal model: hemi and homo α-syn A53T mice exhibiting medium and high dopaminergic neuron loss accompanied by motor deficits from 5 days of sub-toxic (10 mg/kg) injection of MPTP.
Although the exact pathological mechanisms remain unclear, abnormal aggregation and deposition of α-syn have been pivotally implicated in many studies [32]. Since DJ-1 interacts with α-syn, overexpression of DJ-1 reduces α-syn dimerization and α-syn-dependent cellular toxicity is reversed by DJ-1 [33]. In this study, we demonstrated that increased mutant A53T α-syn expression level negatively correlates with the expression of DJ-1 in hemi and homo α-syn Tg mice and SH-SY5Y cells; as mutant A53T α-syn increases, the level of DJ-1 is significantly decreased. Together, these data indicate that overexpression of mutant A53T α-syn may cause the decrease in DJ-1 expression, which may be involved in the increased dopamine cell death of A53T α-syn Tg mice by sub-toxic dose of MPTP treatment. This insight underscores the need to find the possible mechanism through which α-syn aggregation in the synapse decreases DJ-1 expression, subsequently undermining neuroprotective effect which then leads to neurodegenerative disease.
Two elimination pathways are mainly involved to clear α-syn accumulation. One is ubiquitin proteasome system (UPS) and the other is autophagy lysosome pathway (ALP) [34]. Dysfunction in these two systems are associated with the pathogeneses of neurodegenerative diseases [3536]. Inhibition of UPS has been suggested as one of the mechanisms leading to the accumulation of α-syn [37]. UPS involves three enzymes: ubiquitin activating (E1), ubiquitin conjugating (E2) and ubiquitin ligating (E3) [3839]. In our study, the expression level of ubiquitin conjugating enzyme, UBE2N, significantly was decreased in mutant α-syn expressing Tg mice but not in SH-SY5Y cells (data not shown). Our findings suggest that the reduction of DJ-1 may be involved with the impaired activity of proteasome in α-syn Tg mice.
CONCLUSION
Taken together, in the present study we demonstrated that according to the expression level of mutant α-syn in brain, vulnerability of neuronal cells to MPTP or 6-OHDA and behavior impairments were also increased. As a mechanism of that, we observed decreased expression levels of DJ-1, which is known to reduce α-syn aggregation and cellular toxicity, in SN of mutant α-syn expressing Tg mice. These suggest that mutant α-syn expressing Tg may be vulnerable to oxidative injury through reduced DJ-1.



 XML Download
XML Download