

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Osteoarthritis is the most common degenerative articular disease and affects multimillions of people, especially in the elderly. Among the symptoms of osteoarthritis, degeneration of articular cartilage, formation of osteophytes, synovial inflammation, and changes in subchondral bone are the major pathophysiologic features. The cause of osteoarthritis is still unclear, and involves multiple biochemical and mechanical factors. The equilibrium between physiologic synthesis and degradation of articular cartilage is disrupted during the progression of osteoarthritis [12].
In articular cartilage, the activation of degradative enzymes leads to the loss and degradation of proteoglycans and collagen. In particular, the matrix metalloproteinases (MMPs) play a pivotal role in the destruction of articular cartilage in osteoarthritis patients [34]. The types of MMP include collagenases (MMP-1, -8 and -13), gelatinases (MMP-2 and -9), and stromelysins (MMP-3, -7, -10 and -11) [56]. Among them, MMP-3 degrades proteoglycans and activates procollagenase in articular cartilage [78]. In addition to MMP-3, MMP-1 and MMP-13 were reported to play important roles in the destruction of cartilage in osteoarthritis. MMP-1 is a commonly detected metalloproteinase in synovial fluid from patients suffering from osteoarthritis [9101112131415]. Another degradative enzyme, ADAMTS-4, is a major aggrecanase in cartilage of mouse and ADAMTS-5 has been known to be important in cartilage matrix destruction during osteoarthritis [1617].
Therefore, we suggest it is valuable to find the potential activity of regulating (inhibiting) the expression and activity of MMPs by the compounds derived from various medicinal plants used as arthritis remedies in folk medicine, for development of new therapeutic strategies for osteoarthritis. We have tried to investigate the potential activity of some natural products on the expression and activity of MMP-3 in articular chondrocytes. As a result of our trial, we previously reported that several natural compounds affected the gene expression, secretion (production) and proteolytic activity of MMP-3, in vitro and in vivo [18192021].
According to a number of reports, betulin, a natural product derived from Betulae Cortex, a medicinal plant used for controlling various inflammatory diseases in folk medicine, showed the biological activities including anti-inflammatory effect [2223242526]. Betulin showed an alleviating effect on alcoholic liver injury and renal injury through SIRT1/AMPK and TLR4/NF-κB signaling pathways, respectively [2223]. It also attenuates lung injury in sepsis and mammary gland inflammatory injury [2425]. Betulin suppresses the expression of pro-inflammatory cytokines in human cardiac cells [26].
However, to the best of our knowledge, there are no report about the effect of betulin on the gene expression, secretion, and enzyme activity of MMP-3, an articular cartilage-degradative enzyme that decomposes proteoglycans, in primary cultured rabbit articular chondrocytes, or on in vivo production of MMP-3 in the rat knee joint. Therefore, to evaluate the chondroprotective potential of betulin, we investigated its effects on IL-1β-induced gene expression, secretion, and enzyme activity of MMP-3 in vitro, and on production of MMP-3 in vivo.
METHODS
Materials
All chemicals and reagents used in this experiment, including betulin (purity: 98.0%), were purchased from Sigma-Aldrich (St. Louis, MO, USA) unless otherwise specified. Dulbecco's Modified Eagle's Medium (DMEM) was purchased from Gibco-BRL (Grand Island, NY, USA) and recombinant human IL-1β was purchased from R&D Systems (Minneapolis, MN, USA).
Primary cultures of chondrocytes from rabbit articular cartilage
Male New Zealand White rabbits were obtained from Daehan Biolink (Seoul, South Korea) at 2 weeks of age. Animals were housed 1 animal per cage, provided with distilled water and food ad libitum, and kept under a 12 h light/dark cycle (lights on from 08:00~20:00) at constant temperature (22.5℃) and humidity (55%). Animals were cared for in accordance with the Guide for the Care and Use of Laboratory Animals, and care was regulated by Chungnam National University (the approval number of animal experiment: CNU-00555) (Daejeon, Korea). Rabbit articular chondrocytes were isolated from the tibial plateau and femoral condyle in cartilage of the knee joint. Cartilage was washed in phosphate-buffered saline (PBS) and minced into pieces measuring 2 mm3, approximately. Cartilage tissue was digested for 4 h with 0.2% type II collagenase at 37℃. After collection of individual cells by brief centrifugation, the cells were transferred to 100 mm culture dishes (seeding density: 105 cells/cm2) in 12 mL DMEM supplemented with 10% fetal bovine serum (FBS), in the presence of penicillin (100 units/mL) and streptomycin (100 µg/mL). Cells were cultured at 37℃ in a humidified, 5% CO2/95% air, water-jacketed incubator, and medium was replaced every other day [27].
Treatment of cells with betulin
Chondrocytes were seeded on 6-well culture plates (for RT-PCR) or 60 mm culture dishes (for western blotting) at a density of 105 cells/cm2. After 2 days in monolayer culture, the cells were incubated for 2 h in growth medium with 1, 10, 50, or 100 µM of betulin followed by incubation in the presence or absence of IL-1β (10 ng/mL) for 24 h. Betulin was dissolved in dimethylsulfoxide, diluted in PBS, and administered in culture medium (final concentrations of dimethylsulfoxide were 0.5%). The final pH values of these solutions were between 7.0 and 7.4. Culture medium and 0.5% dimethylsulfoxide in medium did not affect the gene expression, secretion, or proteolytic activity of MMP-3 in primary cultured chondrocytes. The supernatant was collected and centrifuged, and cell and supernatant fractions were stored at –80℃ until use.
Cytotoxicity assay
Chondrocytes were seeded at a density of 2×105/mL (0.1 mL/well) in a 96-well microtiter plate, and allowed to attach for 24 h to keep the log phase growth at the time of drug treatment. Betulin was dissolved in DMSO, and administered in DMEM supplemented with 10% FBS (final concentrations of DMSO were under 0.5%). 0.5% DMSO alone did not affect the proliferation of chondrocytes. After incubation with the indicated drug concentrations for 72 h, cell proliferation was determined using the sulforhodamine B (SRB) assay [28].
Isolation of total RNA and RT-PCR
Total RNA was isolated from chondrocytes using the Easy-BLUE Extraction Kit (INTRON Biotechnology, Inc. Kyung-ki-do, South Korea), and reverse transcribed using AccuPower RT Premix (BIONEER Corporation, Daejeon, South Korea) according to the manufacturer's instructions. About 2 µg of total RNA was primed with 1 µg of oligo (dT) in a final reaction volume of 30 µL. 2 µL of RT reaction product was amplified in 20 µL using Thermoprime Plus DNA Polymerase (ABgene, Rochester, NY, USA). PCR was performed with the following primers: MMP-3 (5′ATG GAC CTT CTT CAG CAA 3′, 5′TCA TTA TGT CAG CCT CTC 3′), MMP-13 (5′AGG AGC ATG GCG ACT TCT AC 3′, 5′TAA AAA CAG CTC CGC ATC AA 3′), MMP-1 (5′TCA GTT CGT CCT CAC TCC AG 3′, 5′TTG GTC CAC CTG TCA TCT TC 3′), ADAMTS-4 (5′CAA GGT CCC ATG TGC AAC GT 3′, 5′CAT CTG CCA CCA CCA GTG TCT 3′), ADAMTS-5 (5′TGT CCT GCCAGC GGATGT 3′; 5′ACG GAA TTA CTG TAC GGC CTA CA 3′), and type II collagen (5′AAC ACT GCC AAC GTC CAG AT 3′, 5′CTG ACG CAC GGT ATA GGT GA 3′). GAPDH (5′ACT GGC GTC TTC ACC ACC AT 3′; 5′AAG GCC ATG CCA GTG AGC TT 3′) was used as a quantitative control. The PCR products increased as the concentration of RNA increased. The amplified fragment sizes were 350 base pairs (bp) for MMP-3, 458 bp for MMP-13, 300 bp for MMP-1, 90 bp for ADAMTS-4, 110 bp for ADAMTS-5, 220 bp for type II collagen, and 400 bp for GAPDH. After PCR, 15 µL of PCR products were subjected to 2% agarose gel electrophoresis and visualized with ethidium bromide under a transilluminator [27]. The signal intensity of each band was analyzed by GelQuant software (DNR Bio-Imaging Systems Ltd., Jerusalem, Israel).
Western blot analysis for measuring secretion level of MMP-3 in culture supernatant
Chondrocytes (confluent in 60 mm culture dish) were incubated for 2 h in growth medium with 1, 10, 50, or 100 µM of betulin followed by incubation in the presence or absence of IL-1β (10 ng/mL) for 24 h. After the treatment, the supernatant was collected and the cells were harvested using 3 x trypsin-EDTA solution and then centrifuged in a microcentrifuge (1,200 rpm, 3 min, 4℃). The Bradford assay was used to measure protein concentrations in culture supernatants to ensure consistent weight of protein samples subjected to electrophoresis. Culture supernatant samples containing MMP-3 protein (50 µg each) were subjected to 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE), and then transferred onto a polyvinylidene difluoride (PVDF) membrane. Blots were blocked using 5% skim milk in Tris-buffered saline/Tween 20 (TBS-T), and probed overnight with MMP-3 antibody in blocking buffer at 4℃. Antibody against MMP-3 was purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA). Membranes were washed with TBS-T and probed for 1 h with a secondary antibody conjugated with horseradish peroxidase (Calbiochem, La Jolla, CA, USA). After 4 washes with TBS-T, immunoreactive bands were detected using an enhanced chemiluminescence kit (Pierce ECL western blotting substrate, Thermo Scientific, Waltham, MA, USA). The signal intensity of each band was analyzed by GelQuant software (DNR Bio-Imaging Systems Ltd., Jerusalem, Israel).
Casein zymography to measure the proteolytic activity of MMP-3
A modified casein-substrate zymography was carried out using culture supernatants from chondrocytes pretreated for 2 h with betulin and stimulated for 24 h with IL-1β in DMEM containing 0.5% FBS. The Bradford assay was used to measure protein concentrations in culture media to ensure consistency across samples. Samples were electrophoresed at 4℃ in a 10% SDS gel containing 0.1% casein. After electrophoresis, gels were washed with 10 mM Tris-HCl (pH 8.0) containing 2.5% Triton X-100. Next, gels were incubated at 37℃ for 48 h in 50 mM Tris-HCl (pH 8.0) containing 1% Triton X-100, 0.2 M NaCl, and 5 mM CaCl2. Finally, gels were stained with 1% Coomassie Brilliant Blue, destained, and photographs were taken [27]. The signal intensity of each band was analyzed by GelQuant software (DNR Bio-Imaging Systems Ltd., Jerusalem, Israel).
In vivo experiments
Male Sprague–Dawley rats (Daehan Biolink, Seoul, South Korea) weighing 200~210 g were used to investigate the effect of betulin on production of MMP-3 in articular cartilage in vivo. Animals were housed 5 per cage, provided with distilled water and food ad libitum, and kept under a 12 h light/dark cycle (lights on from 08:00~20:00) at constant temperature (22.5℃) and humidity (55%). Animals were cared for in accordance with the Guide for the Care and Use of Laboratory Animals, and care was regulated by Chungnam National University (the approval number of animal experiment: CNU-00555) (Daejeon, South Korea). Rats were randomly divided into 4 groups: control, IL-1β only, 50 µM betulin plus IL-1β, or 100 µM betulin plus IL-1β. Rats were anesthetized with vaporized diethyl ether, and those from the 50 µM betulin plus IL-1β and 100 µM betulin plus IL-1β treatment groups received a 30 µL injection of 50 µM or 100 µM betulin, respectively, into the right knee joint. After 3 h, rats from the IL-1β only group, the 50 µM betulin plus IL-1β group, and the 100 µM betulin plus IL-1β group received a 30 µL injection of 20 ng IL-1β in sterile PBS into the right knee joint. Rats from the control group were injected with 30 µL of sterile PBS. Rats were euthanized via CO2 asphyxiation 72 h after injections. Articular cartilage (tibial plateau and femoral condyle) was isolated from each animal, homogenized, and prepared for measurement of MMP-3 protein by western blot analysis. Tissue lysates from articular cartilage homogenates containing MMP-3 protein (50 µg each) were subjected to 10% SDS-PAGE, and transferred onto a PVDF membrane. Blots were blocked with 5% skim milk in TBS-T, and probed with MMP-3 antibody (Santa Cruz Biotechnology, Santa Cruz, CA, USA) in blocking buffer overnight at 4℃. Membranes were washed with TBS-T, and probed for 1 h with a secondary antibody conjugated with horseradish peroxidase (Calbiochem, La Jolla, CA, USA). After 4 washes with TBS-T, immunoreactive bands were detected using an enhanced chemiluminescence kit (Pierce ECL western blotting substrate, Thermo Scientific, Waltham, MA, USA). The signal intensity of each band was analyzed by GelQuant software (DNR Bio-Imaging Systems Ltd., Jerusalem, Israel).
RESULTS
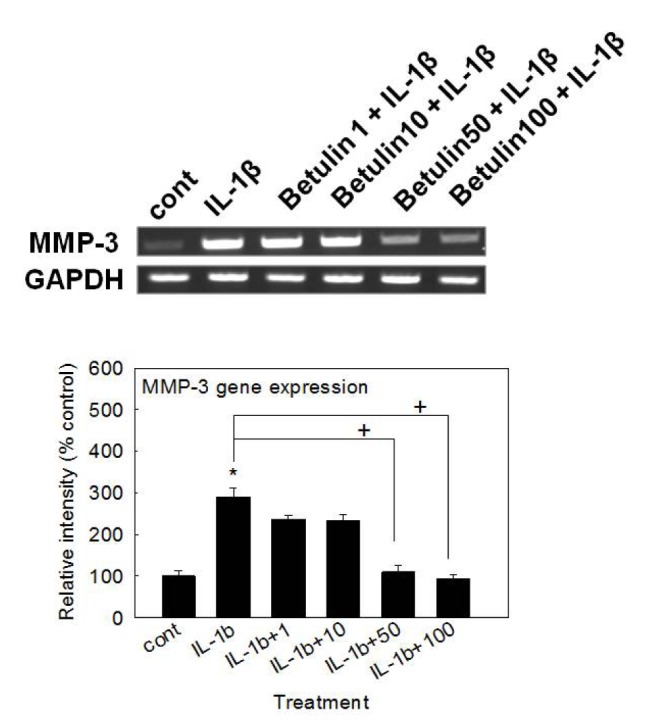
Effect of betulin on MMP-3 gene expression in rabbit chondrocytes
To examine the potential activity of betulin on the gene expression of MMP-3, the key matrix metalloproteinase involved in destruction of articular cartilage, MMP-3 gene expression was measured after pretreatment of betulin. As shown in Fig. 1, betulin inhibited IL-1β-induced MMP-3 gene expression.
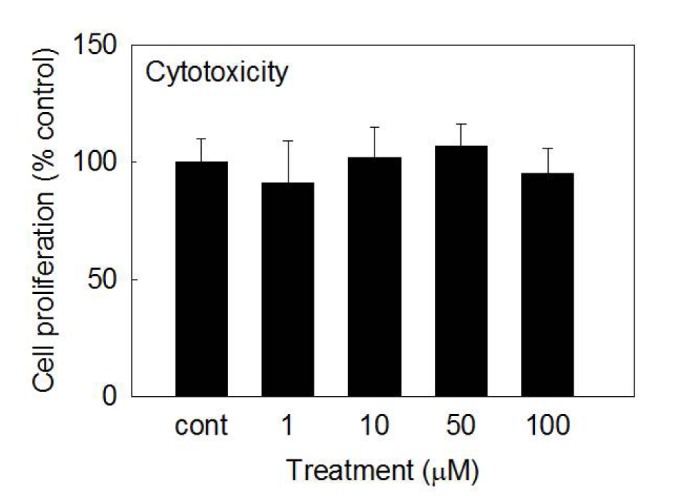
Cytotoxicity of betulin to rabbit chondrocytes
To investigate the potential cytotoxicity of betulin to cultured rabbit chondrocytes, effect of betulin on proliferation of rabbit chondrocytes using SRB assay was tested. As can be seen in Fig. 2, betulin showed no significant cytotoxicity at the concentrations of 1, 10, 50, and 100 µM. The numbers of cells in betulin-treated cultures were 100±10%, 91±18%, 102±13%, 107±9%, and 95±11% for control, 1, 10, 50, and 100 µM betulin, respectively.
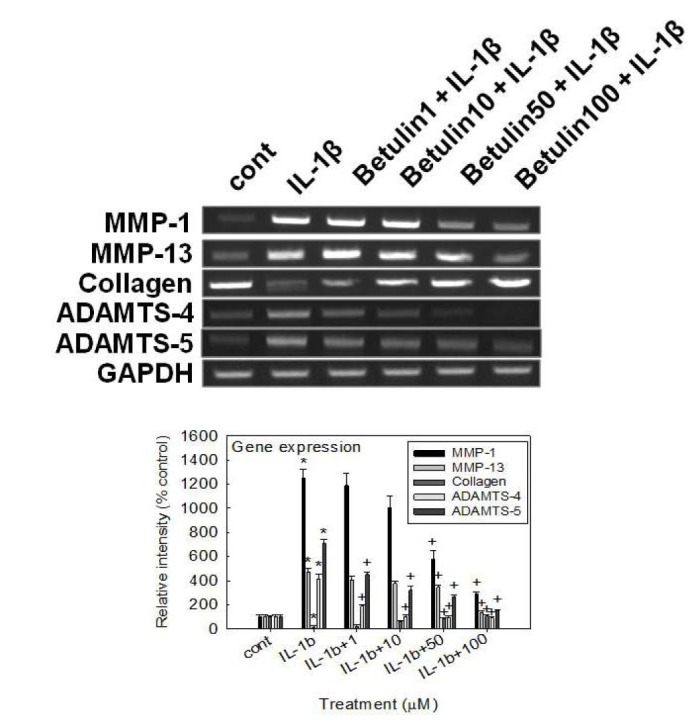
Effect of betulin on the gene expression of MMP-1, MMP-13, ADAMTS-4, ADAMTS-5 or type II collagen in rabbit chondrocytes
If betulin can affect the gene expression of MMP-3, the key matrix metalloproteinase involved in destruction of articular cartilage, it should be investigated whether betulin affects the gene expression of MMP-1, MMP-13, ADAMTS-4 or ADAMTS-5, the other degradative enzymes related to destruction of articular cartilage, and type II collagen, in rabbit chondrocytes. As shown in Fig. 3, betulin showed the suppression of IL-1β-induced gene expression of MMP-1, MMP-13, ADAMTS-4, and ADAMTS-5, in rabbit chondrocytes. Furthermore, betulin showed an additional chondroprotective effect by restoring the compromised gene expression of type II collagen by IL-1β, in rabbit chondrocytes.
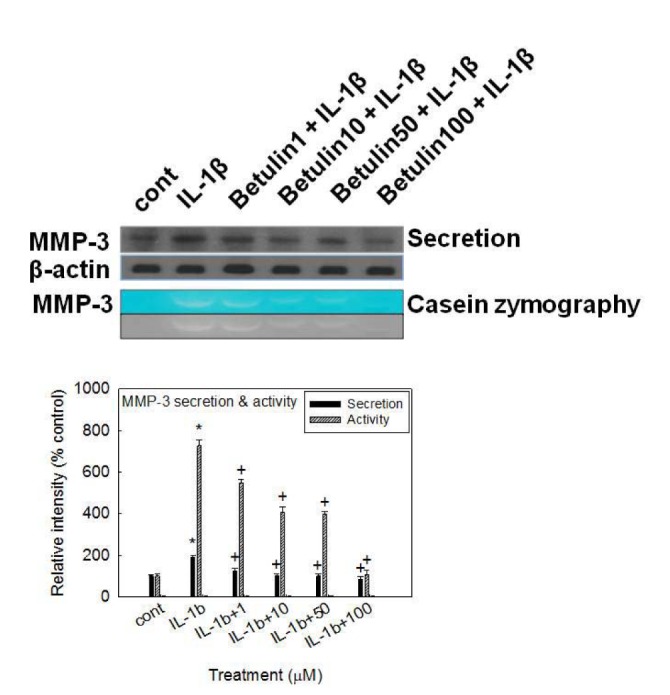
Effect of betulin on IL-1β-induced secretion of MMP-3 from rabbit articular chondrocytes
If betulin can affect the MMP-3 gene expression at the transcriptional level, it should be investigated whether betulin affects IL-1β-induced secretion of MMP-3 proteins from rabbit articular chondrocytes. As shown in Fig. 4, stimulation with IL-1β (10 ng/mL) increased secretion of MMP-3 from chondrocytes. However, betulin inhibited the effects of IL-1β on MMP-3 secretion.
Effect of betulin on proteolytic activity of MMP-3 in rabbit articular chondrocytes
To investigate the effect of betulin on the enzyme activity of secreted MMP-3, which is known to degrade proteoglycans, one of the two major matrix components of cartilage, culture supernatants from rabbit articular chondrocytes were analyzed for caseinolytic activity by casein zymography, after treatment with IL-1β for 24 h. As can be seen in Fig. 4, IL-1β increased the caseinolytic activity of MMP-3 in rabbit articular chondrocytes, and this effect was inhibited by pretreatment with betulin.
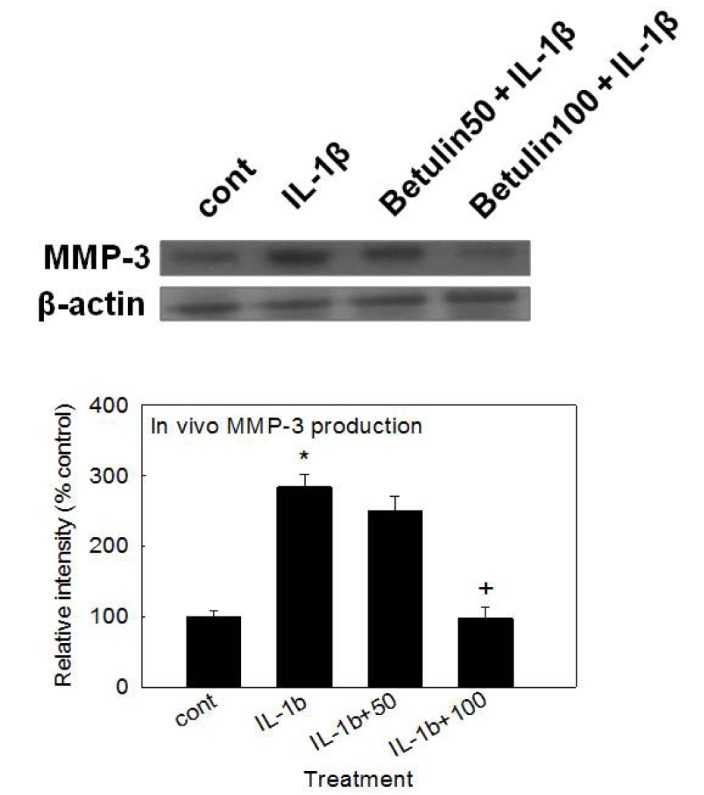
Effect of betulin on MMP-3 production in vivo
To investigate whether betulin shows the potential effect in vivo, we examined the effect of intraarticular injection of betulin into the knee joint of rats on IL-1β-stimulated production of MMP-3 from articular cartilage tissues. As shown in Fig. 5, treatment with IL-1β (20 ng/30 µL) increased MMP-3 production in articular cartilage tissues. However, betulin inhibited IL-1β-induced MMP-3 production, in vivo.
DISCUSSION
To restore the broken equilibrium between physiological synthesis and degradation of articular cartilage during the progression of osteoarthritis, discovering a useful and specific pharmacological tool can be a promising approach to the effective control of this condition. Although osteoarthritis can be defined as a non-inflammatory disease, its development and progression have been attributed to low-grade inflammation in intraarticular sites, as well as to various inflammatory cytokines in articular tissues and fluids that are produced by chondrocytes and/or interact with chondrocytes [29303132].
IL-1β is an inflammatory cytokine that is produced by cells in articular tissues, including chondrocytes, and which can increase expression of MMPs and stimulate the progression of osteoarthritis. IL-1β plays an important role in the initiation and progression of destruction of articular cartilage by suppressing synthesis of collagen and stimulating MMP expression [313334]. Particularly, MMP-3 has been reported to play a pivotal pathophysiological role in osteoarthritis by degrading components of the extracellular matrix, such as proteoglycans. MMP-3 levels were increased more than MMP-1 levels in patients suffering from osteoarthritis in knee joints compared to the control group [735].
According to several reports, IL-1β-stimulated expression of MMPs is associated with the suppression of the NF-κB signaling pathway [36373839]. Jung and his colleagues reported that Schisandrae Fructus, an anti-inflammatory medicinal plant used in folk medicine, inhibited IL-1β-induced expression and activity of MMP-1, MMP-3, and MMP-13 and markedly suppressed the nuclear translocation of NF-κB by blocking IkB-alpha degradation in SW1353 human chondrocytes [36]. Taraxasterol, a natural product with anti-inflammatory effect, suppressed IL-1β-induced expression of MMP-1, MMP-3, MMP-13 and activation of NF-κB [37]. Another natural product, matrine, inhibited IL-1β-induced expression of MMPs by suppressing the activation NF-κB in human chondrocytes in vitro [38]. Zhou et al. reported that coptisine, an isoquinoline alkaloid extracted from an antiinflammatory medicinal plant, also inhibited the expression of MMP-3 and MMP-13 through inhibition of NF-κB activation in human chondrocytes [39].
IL-1β activates several intracellular signal transduction cascades among which the NF-κB pathway is pivotal. NF-κB is a heterodimer composed of p65, p50 and IkBα subunits present in the cytoplasm as an inactive state. In response to various stimuli, the IkBα subunit is phosphorylated and degraded, thereby facilitating the translocation of p50-p65 heterodimer to the nucleus. The p50-p65 acts as a transcription factor regulating the expression of numerous genes including MMP-3 [36]. In the present study, betulin affected the gene expression, secretion, and proteolytic activity of MMP-3, by directly acting on articular chondrocytes. We found that betulin inhibited IL-1β-induced gene expression of MMP-3, MMP-1, MMP-13, ADAMTS-4, and ADAMTS-5 and restored the gene expression of type II collagen that had been inhibited by IL-1β, in rabbit articular chondrocytes (Fig. 1 and 3). Thus, the chondroprotective effect of betulin are supported by its regulation of the gene expression of diverse proteases involved in the destruction of articular cartilage in osteoarthritis, as well as by its promotion of the gene expression of type II collagen at the transcriptional level. Additionally, IL-1β-stimulated secretion and the proteolytic activity of MMP-3 from articular chondrocytes were suppressed by betulin (Fig. 4). This result means that betulin can regulate the step of protein synthesis and secretion of MMP-3 and affects the proteolytic activity of overproduced and oversecreted MMP-3 in tissues of osteoarthritic articular cartilage.
The underlying mechanism of action of betulin on the gene expression, secretion, and proteolytic activity of MMP-3 are not clear at present, although we are investigating whether betulin act as a potential regulator of NF-κB signaling pathway in articualr chondrocytes, based on a report on the inhibitory effect of betulin on lipopolysaccharide (LPS)-stimulated rat HBZY-1 renal mesangial cells through inactivation of NF-κB signaling pathway [23]. Lastly, we investigated the effect of intraarticular injection of betulin into the knee joint of rats on IL-1β-stimulated production of MMP-3 in articular cartilage tissue. As can be seen in Fig. 5, betulin inhibited IL-1β-stimulated production of MMP-3 in articular cartilage tissue. This result shows that, in addition to its in vitro effects, betulin exerts chondroprotective effects in vivo when administered via intraarticular injection.
Taken together, the inhibitory action of betulin on the gene expression, secretion, and enzyme activity of MMP-3 in articular chondrocytes and production of MMP-3 in the knee joint of rats might explain, at least in part, the traditional use of Betulae Cortex as an anti-inflammatory agent for diverse inflammatory diseases, in folk medicine. We suggest it is valuable to find the natural products that have specific suppressive effects on the gene expression, secretion, and enzyme activity of MMP-3 - in view of both basic and clinical sciences - and the result from this study suggests a possibility of developing betulin as a candidate for novel agent controlling cartilage damage in osteoarthritis via intraarticular administration, although further studies are essentially required.
 XML Download
XML Download