

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
The transient receptor potential (TRP) channel was initially cloned in fruit flies [1]. The TRP channel is activated by rhodopsin, a G protein-coupled receptor (GPCR), and responds to light. Since the initial reports, GPCR-activated TRP channels were subsequently cloned in mammalian tissues and named classical type TRP (TRPC) channels. The TRPC subfamily consists of seven known members [2]. Some studies have shown that TRPCs 1, 3, 4, and 5 can be activated by depletion of endoplasmic reticulum Ca2+ stores, suggesting that these channels contribute to SOC entry. On the contrary, other studies have proposed that these homologues contribute to ROC entry [3]. In the studies that focused specifically on endogenous TRPC proteins contributing to SOC entry in endothelial cells, TRPC1 and TRPC4 were shown to contribute to the endothelial Ca2+-selective ISOC channel. Previously, antisense technology was used to assess the contribution of endogenous TRPC1 to ISOC in pulmonary artery endothelial cells (PAECs) [4]. Inhibition of TRPC1 reduced but did not eliminate ISOC. On the other hand, endothelial cells obtained from TRPC4-deficient mice completely lacked ISOC [5]. These studies suggest that, although TRPC1 and TRPC4 both contribute subunits to the ISOC channel, only TRPC4 is required for channel activation.
TRP channels are calcium-permeable cation channels. As a family member of TRP channel, TRPC channels are nonselective cation channels permeable to calcium as well as monovalent cations. When calcium permeability is calculated with shift of reversal potentials, the permeability of these channels to monovalent cations ranges from 1 to 7 [678]. On the other hand, TRPC4 [a component of the Ca2+ release-activated Ca2+ (CRAC) channel] has been calculated to have a higher ratio of 159.7 [5]. The permeability ratio has been shown to differ depending on whether the I-V curve has a doubly rectifying shape or an inward rectifying shape.
In gastrointestinal smooth muscle, acetylcholine binds to the muscarinic receptor and induces calcium release from calcium stores through the inositol triphosphate (IP3) receptor [9]. In addition, acetylcholine activates nonselective cation channels and induces membrane depolarization, which in turn activates the voltage-operated calcium channel (VOCC). The mobilization of calcium through the VOCC increases the intracellular calcium. TRPC4 channels are candidate nonselective cation channels that are activated by muscarinic stimulation and contribute to membrane depolarization rather than to intracellular calcium increase [10].
Calcium ions modulate the TRPC5 channel through direct action, calmodulin [1112], and myosin light chain kinase (MLCK) [1314]. TRPC4 belongs to the same subfamily as TRPC5. Schaefer et al. [8] also observed critical dependence of TRPC4 and TRPC5 activity on [Ca2+]i. Calcium influx through the TRPC4 channel might modulate TRPC channels near the channel itself as a feedback mechanism [15].
GCaMP is one of the genetically encoded calcium indicators (GECIs) which is a powerful tool for tracking the Ca2+ dynamics in target cells. GCaMP is composed of green fluorescent protein (GFP), calmodulin (CaM) and CaM-interacting M13 peptide. Ca2+ binding to the CaM result in a conformational change in CaM-M13 complex, eliminates bright fluorescence. Previously, a GCaMP3-tagged TRPML channel was used to measure lysosomal calcium release [16]. GCaMP3-TRPML1 responded to calcium released from the lysosome, but not to calcium released from the endoplasmic reticulum (ER), indicating the existence of calcium nanodomains near the lysosome. To measure the calcium concentrations near specific channels, we tagged channels at their C-terminus with GCaMP6s. Using this construct, we investigated the calcium permeability of the TRPC4 channel, the role of calcium in TRPC4 function, and the role of TRPC1 in the calcium permeability of TRPC4 channels.
METHODS
Cell culture and transfection
Human embryonic kidney (HEK293) cells were incubated in Dulbecco's Modified Eagle's Medium (DMEM) supplemented with 10% heat-inactivated FBS and penicillin (100 units/ml), streptomycin (100 µg/ml) at 37℃ in 5% CO2 humidified incubator. The mTRPC4β-GCaMP6s construct was made by inserting the full-length GCaMP6s sequence between the AgeI and NotI site of a pEYFP-N1 plasmid which contains the mouse TRPC4β complementary DNA. All constructs were confirmed by sequencing. Every plasmid DNA was prepared using a plasmid midi kit (QIAGEN) as detailed in the protocol. For the transfection, the cells were seeded in 12-well plates. The following day, mTRPC4β-GCaMP6s and other genetically encoded calcium indicators (GECI) were also transfected for electrophysiology and Ca2+ imaging by using the transfection reagent FuGENE 6 (Roche Molecular Biochemicals), as detailed in the manufacture's protocol. We co-expressed mTRPC4β-GCaMP6s and type 2 muscarinic (M2) receptor by 1:1, mTRPC4β-GCaMP6s and hTRPC1α by 2:1 ratio.
Solutions and drugs
The patch pipette containing standard intracellular solution; 140 mM CsCl, 10 mM HEPES, 0.5 mM EGTA, 3 mM Mg-ATP, 0.2 mM Tris-GTP, PH 7.3 with CsOH. External solution was perfused constantly as follows; 3.6 mM KCl, 10 mM HEPES, 1 mM MgCl2, 145 mM NaCl, 2 mM CaCl2, 5 mM glucose, PH 7.4 with NaOH. We used TRPC4 channel agonist, (–)-englerin A (EA, Sigma-Aldrich) and muscarinic receptor agonist, carbachol (CCh, Sigma-Aldrich). To identify the only effect of CCh to TRPC4 channel, we changed solution from normal tyrode (NT) to thapsigargin (TG, Sigma-Aldrich) first to block the ability to pump calcium into the ER. We used Ca2+ ionophore, ionomycin (IM, Sigma-Aldrich) to raise the intracellular calcium level.
Electrophysiological recordings
The cells were transferred onto a small chamber on the stage of an inverted microscope (I×70, Olympus, Japan) and attached to coverslip in the small chamber for 10 min prior for the patch recording. Glass microelectrodes with 2~2.5 megaohms resistance were used to obtain gigaohm seals. The currents were recorded using an Axopatch 200B patch-clamp amplifier (Axon instrument, USA). The whole cell configuration was used to measure the TRPC4 channel current in the HEK293 cells. Voltage ramps ranging from +100 to –100 mV over period of 500 msec were imposed every 10 sec with a holding membrane potential of –60 mV. pCLAMP software (version 10.2, Axon instrument, USA) were used for data acquisition and the data were analyzed using the OriginPro 8 (OriginLab, USA). The recording chamber was continuously perfused at a flow rate of 1~2 ml/min.
Calcium imaging
To obtain the image to measure calcium influx, we used an inverted microscope (I×70, Olympus, Japan) with 60× oil objective lens. Each image was captured on EMCCD camera (iXon3, Andor, Northern Ireland) every 2 sec and the fluorescence intensity at 500 nm (F500) was monitored using the MetaMorph7.6 software (Molecular devices, USA) system. Whole cell current and calcium imaging were recorded simultaneously in almost all experiments.
FRET efficiency computation for YC6.1 [17]
The FRET ratio (FR) is equal to the fractional increase in YFP emission due to FRET and was calculated as FR=FAD
AD/FA=[SFRET(DA)–RD1·SCFP (DA)]/(RA1·[SYFP (DA)–RD2·SCFP (DA)]). Here, SCUBE(SPECIMENDA) denotes an intensity measurement, where CUBE indicates the filter cube (CFP, YFP, or FRET), and SPECIMEN indicates whether the cell is expressing the donor (D; CFP), acceptor (A; YFP), or both (DA). RD1=SFRET(D)/SCFP(D), RD2=SYFP(D)/SCFP(D), and RA1=SFRET(A)/SYFP(A) are predetermined constants from measurements applied to single cells expressing only CFP- or YFP-tagged molecules. Although three-cube FRET does not require that CFP and YFP fusion constructs preserve the spectral features of the unattached fluorophores, similar ratios and recorded spectra furnished two indications that the spectral features of the fluorophores were largely unperturbed by fusion. Since the FR relies on YFP emission, YFP should be attached to the presumed limiting moiety in a given interaction. Subsequent quantitative calculations based on FR relied on a presumed 1:1 interaction stoichiometry. The effective FRET efficiency (EEFF) was determined by EEFF=E·Ab=(FR–1)·[EYFP(440)/ECFP(440)], where E is the intrinsic FRET efficiency when fluorophore-tagged molecules are associated with each other, Ab is the fraction of YFP-tagged molecules that are associated with CFP-tagged molecules, and the bracketed term is the ratio of YFP and CFP molar extinction coefficients scaled for the FRET cube excitation filter [18]. We determined this ratio to be 0.094 based on maximal extinction coefficients for ECFP and EYFP [19] and excitation spectra.
RESULTS
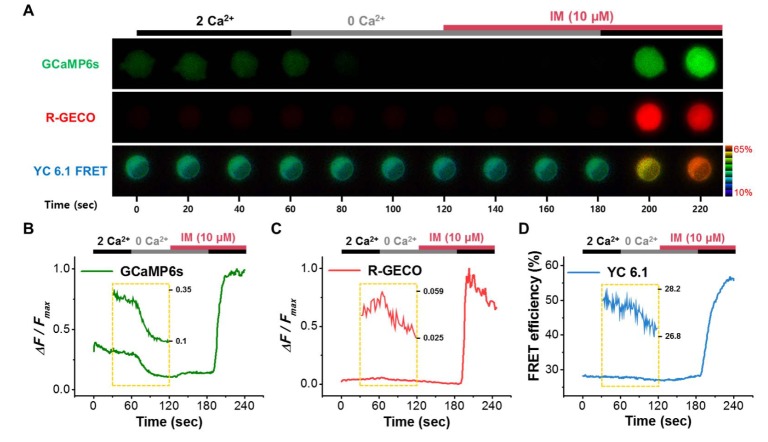
Three GECIs were used to detect calcium changes, GCaMP6s, R-GECO, and Yellow Cameleon 6.1 (YC 6.1) (Fig. 1). Depending on Kd value, the GECIS exhibited different responses to changes in extracellular calcium and ionomycin (IM, 10 µM) (Fig. 1A). GCaMP6s responded to a change in external calcium from 2 mM to 0 mM (Fig. 1B). On the other hand, R-GECO did not respond to this change (Fig. 1C). YC6.1 has a similar Kd value to GCaMP6s, and responded to calcium addition induced by IM, but with slower kinetics than GCaMP6s (Fig. 1D). Based on these results, we tagged the C-terminus of the mTRPC4β channel with GCaMP6s.
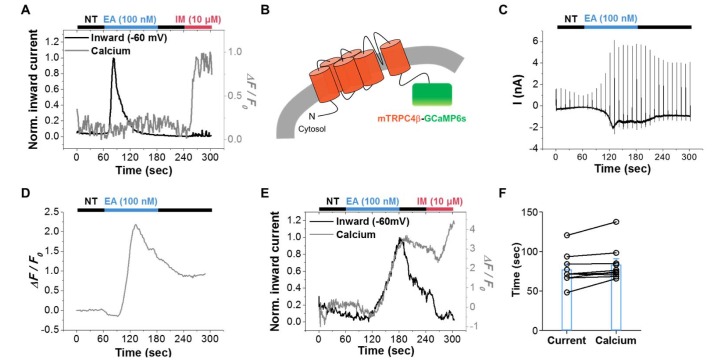
We first measured current and calcium signal simultaneously and compared their values with those from cells expressing both mTRPC4β and GCaMP6s (Fig. 2). Englerin A (EA, 100 nM) [2021], a new agonist of TRPC4/5 channels, induced a large current but did not increase the calcium signal (Fig. 2A). On the other hand, IM did not induce any change in current but increased the calcium signal. A previous report concluded that TRPC4 is an SOC and contributes to intracellular calcium increase [22]; our data contradict this finding. Our results suggest that cytosolic GCaMP6s do not detect the calcium change induced by mTRPC4β activation, and that mTRPC4β does not contribute a significant amount of calcium to the cytosol. We next attached GCaMP6s to the C-terminus of mTRPC4β, thereby generating an mTRPC4β-GCaMP6s construct, and tested whether this GCaMP6s tag could detect calcium influx through TRPC4 channels (Fig. 2B). Englerin A induced a large current in mTRPC4β-GCaMP6s, similar to its action on TRPC4 channels (Fig. 2C). The I-V curve showed a doubly rectifying shape. The measured calcium signal is shown in Fig. 2D. EA increased the fluorescence intensity of GCaMP6s attached to mTRPC4β. In contrast to free GCaMP6s, GCaMP6s attached to TRPC4 channels could detect calcium changes near the channels. We next measured the current and calcium signals simultaneously (Fig. 2E). EA increased both the current and the calcium fluorescence; these values returned to those of the control after washout of EA. In contrast, IM increased the calcium fluorescence but did not increase the current. The time to the peak current (77.2±6.8 sec, n=9) was not significantly different from the time to peak calcium fluorescence (83.4±7.5 sec, n=9) (Fig. 2F). These results indicate that calcium can permeate the TRPC4 channels and increase calcium concentration near the channels, but not throughout the cytosol. In addition, mTRPC4β-GCaMP6s is a useful tool for testing calcium permeability through TRPC4 channels.
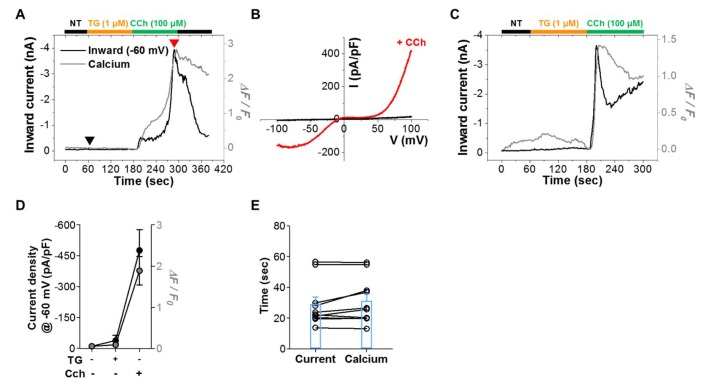
We also investigated whether physiological signaling pathways induce calcium increase through TRPC4 channels (Fig. 3). TRPC4 channels are activated by muscarinic receptor stimulation; thus, we coexpressed the type 2 muscarinic (M2) receptor with mTRPC4β-GCaMP6s. Carbachol (CCh, 100 µM) increased both current and calcium signal with similar time course as EA stimulation (Fig. 3A) showed. The I-V curve showed a typical doubly rectifying shape, consistent with previous reports (Fig. 3B) [2324252627]. The current at -60 mV increased from -10. 6±5.5 pA/pF (n=10) to –476.9±100.4 pA/pF (n=10) (Fig. 3D), while the calcium signal increased from 0.055±0.018 (n=10) to 1.888±0.343 (n=10). The times to peak current (29.1±4.7 sec, n=10) and calcium fluorescence (31.1±4.7 sec, n=10) were both shorter than those in EA stimulation (Fig. 3E). The responses to CCh were of two types. In the first type, the response was rapid and reached its peak around 15 sec (Fig. 3C). In the second type, activation exhibited some delay, and the time to peak was 60 sec (Fig. 3A). The current and calcium signal were well matched, even in this delayed activation phase. These results suggest that Ca2+ influx through the TRPC4 channel either potentiates TRPC4 directly or induces calcium-induced calcium release (CICR), which further activates TRPC4. In situations with the first type of response, CCh effectively induced IP3-mediated Ca2+ release by increasing IP3 level via PIP2 hydrolysis.
We also tested the effect of thapsigargin (TG, 1 µM) on current and calcium signals. TG did not induce a calcium increase in GCaMP6s-tagged TRPC4 (0.090±0.036, n=10) (Fig. 3A). Similarly, TG did not increase the current (–38.6±24.2 pA/pF, n=10). These results suggest that TRPC4 does not act as a SOC, as previously suggested.
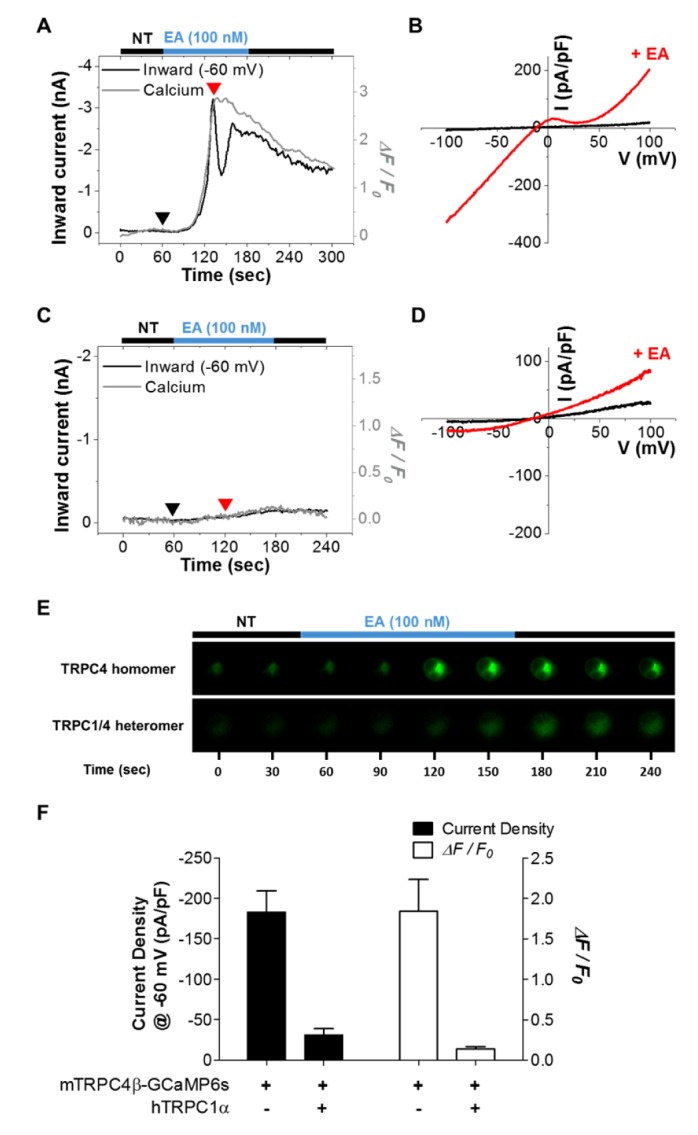
TRPC1 can only mobilize to the plasma membrane when TRPC4 or TRPC5 is coexpressed [2427]. TRPC1 modulates calcium permeability and changes the I-V relationship of the TRPC4 and TRPC5 channels [2428]. To investigate whether TRPC1 affects the detection of calcium near TRPC4 channels, CFP-hTRPC1α was coexpressed with mTRPC4β-GCaMP6s (Fig. 4), and EA was used to activate both TRPC4 homomers and TRPC1/4 heteromers [2021]. EA induced a larger current in TRPC4 homomers (–183.0±26.0 pA/pF, n=10) compared with TRPC1/4 heteromers (–31.2±7.7 pA/pF, n=5) (Fig. 4F). Moreover, the calcium signal increase was greater in TRPC4 homomers (1.84±0.39, n=10) than in TRPC1/4 heteromers (0.141±0.025, n=5). Calcium fluorescence was also visibly decreased in TRPC1/4 heteromers compared to TRPC4 homomers (Fig. 4E). These results indicate that TRPC1 reduces the calcium permeability of TRPC4.
DISCUSSION
In the present study, we showed that 1) mTRPC4β-GCaMP6s functions as a normal TRPC4 channel, despite the presence of the GCaMP6s at the C-terminus. The I-V curve showed a doubly rectifying shape and reversed at 0 mV. 2) The GCaMP6s attached to mTRPC4β detected calcium near TRPC4 channels, but not cytosolic calcium released from the ER. 3) EA and CCh induce current and calcium both at the similar time courses. 4) Coexpression of TRPC1 with TRPC4 induced a smaller calcium increase than that induced by expression of TRPC4 alone.
The time course of calcium increase matched that of current increase. Ca2+ influx through the mTRPC4β channel contributed to the fluorescence change of GCaMP6s near these channels. There are three types of GCaMP6 with different Kd values, GCaMP6s (144 nM), GCaMP6m (167 nM), and GCaMP6f (375 nM). We used GCaMP6s, which has the lowest Kd value and the highest dynamic range (63). Although GCaMP6m has a similar Kd value, the dynamic range (38) was smaller than that of GCaMP6s [29]. The time course of calcium increase exactly matched that of current increase. In some cells (Fig. 3C and Fig. 4A), the calcium signals were maintained longer than the current signals. This finding might reflect the kinetics of the calcium signal compared with those of the current signal.
The TRPC1 channel regulates the calcium permeability of TRPC4 or TRPC5 when they are coexpressed [28]. We found that heteromerization of TRPC4 with TRPC1 decreased GCaMP6s-mediated detection of calcium increase. TRPC1 has been shown to change the I-V curve of TRPC4 and TRPC5 [242728], but not that of TRPC3/6/7 [28]. TRPC1 reduced the calcium permeability of other TRPC channels. The precise reason why TRPC1 has different effects on the I-V curves of the different TRPC channels remains unclear. Further investigations employing TRPC1 and TRPC3/6/7-GCaMP6s would help clarify whether TRPC1 modulates the calcium permeability of TRPC3/6/7.
Typically, TRPC channel functions have been analyzed by calcium addition experiments after the application of 0 mM Ca2+, especially after Ca2+ store depletion with cyclopiazonic acid (CPA) or TG. In this assay, a store-operated calcium channel serves as the source of calcium. Using these assays, TRPC4 and TRPC5 were suggested to be SOC channels [3031]. STIM and ORAI have been suggested to be the main molecules for SOC channels, especially CRAC channels. In calcium addition experiments, the calcium increase was larger when TRPC4 or TRPC5 channels were expressed compared with control conditions. However, even without TRPC4 expression, the calcium addition protocol induced a calcium increase. The expression of TRPC4 resulted in an additional calcium increase above the basal level. We preferred current measurement to calcium measurement for TRPC4 channels, since calcium measurements always include calcium released from CRAC channels as well as from the expressed channels. Thus, GCaMP6s-tagged TRPC4 or TRPC5 channels could be used as tools for studying structure-function relationships by channel calcium permeability.
TRPC5 has been shown to be activated by intracellular calcium [111232]. Moreover, Ca2+ entry via coexpressed CRAC (STIM1 and Orai1) or L-type voltage-operated Ca2+ channels (VOCC) was sufficient to activate TRPC5 channels, suggesting functional interactions between TRPC5 and other Ca2+-selective channels [32]. We found that TRPC4 was activated by intracellular calcium, similar to TRPC5 (Fig. 3A and 3C). TRPC4 is a candidate nonselective cation channel (NSCC) activated by muscarinic stimulation [3334]. Calcium is well known to facilitate the activation of NSCC, which is in turn activated by muscarinic stimulation [10]. Further experiments are required to determine whether the effect of calcium is direct or mediated by CaM.
The interaction of stromal interaction molecule (STIM) and ORAI, two key components of Ca2+ release-activated Ca2+ (CRAC) channels, with TRPC channels in the formation of diverse SOC channels is still a subject of debate. STIM1 binds to and directly regulates the TRPC1, TRPC4, and TRPC5 channels. TRPC5 was specifically examined as a template ROC to clarify the role of STIM1 in ROC regulation. Knockdown of STIM1 with siRNA specifically suppressed CCh-stimulated, but not La3+-stimulated TRPC5 current [35]. A general model for STIMregulated heteromeric Orai/TRPC in SOC/ICRAC channels has been proposed [36]. Orai1 appears to interact with both the TRPC channel N-terminus and C-terminus; although both channel types can function independently of each other [37], there is growing evidence that Orai1 and TRPCs exist in the same Ca2+ signaling complex and influence the activity of each other [38]. When TRPC1 and TRPC4 formed a heteromeric channel, the N-terminal coiled-coil domain (CCD) and C-terminal region (residues 725~745) of TRPC1 were shown to interact with the N-terminal CCD and C-terminal region (residues 700~728) of TRPC4 [27]. However, when TRPC1 and TRPC5 formed a heteromeric channel, the N-terminal CCD and C-terminal region (residues 673~725) of TRPC1 interacted with the N-terminal CCD and C-terminal region (residues 707~735) of TRPC5. Thus, the N-terminal CCD of TRPC channels is essential for the heteromeric structure of TRPC channels, whereas specific C-terminal regions are required for unique heteromerization between subgroups of TRPC channels [27].
In conclusion, GCaMP6s attached to the C-terminus of TRPC4 can detect slight calcium changes at proximal channels. Thus, TRPC4-GCaMP6s can be a useful tool for testing calcium permeability through TRPC4 channels.
 XML Download
XML Download