

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Lim and his coworkers [123] have found that all of TGS, panaxadiol and panaxatriol cause the increased secretion of catecholamines (CA) from the isolated perfused rabbit adrenal glands in a Ca2+-dependent fashion, which are mediated by the activation of cholinergic (both nicotinic and muscarinic) receptors and partly the direct action on the rabbit adrenomedullary chromaffin cells. However, it has been reported that TGS shows the inhibitory effect on the CA release evoked by stimulation of nicotinic receptors in the perfused adrenal medulla of rats [4] and SHRs [5], and also that several ginseng saponins (ginsenosides) from the ginseng root inhibit the CA secretion from bovine adrenal chromaffin cells stimulated by ACh [6789].
Moreover, it has been known that ginsenosides or ginseng extracts cause the hypotensive action [1011121314] while they can rather produces the hypertensive action [15161718]. TGS is found to produce the pressor and depressor actions in the anesthetized normotensive rats [19].
Furthermore, ginseng, when given at small dose in spontaneously hypertensive rat (SHR), causes pressor response, but at relatively large dose rather produces dose-dependent hypotensive response with decreased plasma renin activity [202122]. Sokabe and his coworkers [23] have shown that administration of Korean Red ginseng powder for 11 weeks has no effect on blood pressure in normotensive Donryu (DON) rats, SHR and renal hypertensive rats, whereas it elevates slightly blood pressure in deoxycorticosterone salt hypertensive rats. Recently, it has also been reported that long-term use of North American ginseng had no effect on 24-hour BP and renal function in hypertensive individuals [24].
In the conscious rats, the releasing effect of NO of KRG may be partly contributed to the hypotensive effect of KRG [12]. It has also been shown that Rg3 relaxes the rat thoracic aorta as a consequence of NO production [25]. Han and his co-workers [26] have found evidence that NO levels in exhaled breath of human volunteers by KRG were increased along with decreased blood pressure and heart rate. In a series of studies, it has been found that ginsenosides reduce blood pressure via increases in production of endothelial nitric oxide [10] and that Rg3 is the most potent ginsenoside that activates eNOS in rat aorta [27]. Although some investigators have demonstrated that Rg3 induces eNOS activation in the vasculature of animal models [1028], Hien and his co-workers [29] have found that Rg3 activates eNOS via eNOS phosphorylation in ECV 304 human endothelial cells and increases in eNOS expression. Meanwhile, some studies have shown that ginseng contains a novel G protein-coupled lysophosphatidic acid (LPA) receptor ligand, known as gintonin [303132]. Recently, it has been reported that treatment of gintonin to PC12 cells increased intracellular calcium ([Ca2+]i) as well as the release of dopamine in a concentration-dependent manner [33]. It was concluded that this gintonin-mediated [Ca2+]i elevation is coupled to dopamine release via LPA receptor activation.
Despite of these many studies, there is still controversy about the functional effects of ginsenosides on the cardiovascular system, especially blood pressure and adrenal CA secretion. Therefore, the aim of the present study was to investigate the ability of gintonin, known as a novel G protein-coupled LPA receptor ligand, on the CA secretion in the perfused model of rat adrenal glands, and to establish its mechanism of action.
METHODS
Experimental procedure
All procedures involving animal experiments were conducted in strict accordance with the guidance for the care and use of Laboratory Animals of the National Institutes of Health (NIH Publications No. 80-23; revised in 1996), and approved by the Committee of Experimental Animals, School of Medicine, Chosun University.
Male Sprague-Dawley rats, weighing 180 to 250 grams, were anesthetized with thiopental sodium (40 mg/kg) intraperitoneally. The adrenal gland was isolated by the methods described previously [34]. The abdomen was opened by a midline incision, and the left adrenal gland and surrounding area were exposed by placing three hook retractors. The stomach, intestine and portion of the liver were not removed, but pushed over to the right side and covered by saline-soaked gauge pads and urine in bladder was removed in order to obtain enough working space for tying blood vessels and cannulations.
A cannula, used for perfusion of the adrenal gland, was inserted into the distal end of the renal vein after all branches of adrenal vein (if any), vena cava and aorta were ligated. Heparin (400 IU/ml) was injected into vena cava to prevent blood coagulation before ligating vessels and cannulations. A small slit was made into the adrenal cortex just opposite entrance of adrenal vein. Perfusion of the gland was started, making sure that no leakage was present, and the perfusion fluid escaped only from the slit made in adrenal cortex. Then the adrenal gland, along with ligated blood vessels and the cannula, was carefully removed from the animal and placed on a platform of a leucite chamber. The chamber was continuously circulated with water heated at 37 ±1oC.
Perfusion of adrenal gland
The adrenal glands were perfused by means of ISCO pump (WIZ Co.) at a rate of 0.33 ml/min. The perfusion was carried out with Krebs-bicarbonate solution of following composition (mM): NaCl, 118.4; KCl, 4.7; CaCl2, 2.5; MgCl2, 1.18; NaHCO3, 25; KH2PO4, 1.2; glucose, 11.7. The solution was constantly bubbled with 95% O2+5% CO2 and the final pH of the solution was maintained at 7.4~7.5. The solution contained disodium EDTA (10 µg/ml) and ascorbic acid (100 µg/ml) to prevent oxidation of catecholamine.
Drug administration
Perfusion of gintonin (1~30 µg/ml) for 4 min or single injection of acetylcholine (5.32 mM) in a volume of 0.05 ml were made into perfusion stream via a three way stopcock. In the preliminary experiments it was found that upon administration of the above drugs, secretory responses to gintonin and acetylcholine returned to preinjection level in 4 min. Generally, the adrenal glands were perfused with normal Krebs solution for about one hour before the experimental protocols are initiated.
Collection of perfusate
As a rule, prior to stimulation with gintonin, samples were collected (4 min) to determine the spontaneous secretion of CA ("background sample"). Immediately after the collection of the "background sample", collection of the perfusate was continued in another tube as soon as the perfusion medium containing gintonin reached the adrenal gland. Each perfusate was collected for 4 min at 15 min interval. The amounts secreted in the "background sample" have been subtracted from those from secreted the "stimulated sample" to obtain the net secretion value of CA, which is shown in all of the figures.
To study the effect of a test agent on the spontaneous and drug-evoked secretion, the adrenal gland was perfused with Krebs solution containing the agent for 90 min. Then the perfusate was collected for a specific time period ("background sample"), and then the medium was changed to the one containing the test agent or along with a specific antagonist, and the perfusate were collected for the same period as that for the "background sample". The adrenal perfusate was collected in chilled tubes.
Measurement of catecholamines
CA content of perfusate was measured directly by the fluorometric method of Anton and Sayre [35] without the intermediate purification alumina for the reasons described earlier [34] using fluorospectrophotometer (Kontron Co. Italy).
A volume of 0.2 ml of the perfusate was used for the reaction. The CA content in the perfusate of stimulated glands by secretagogues used in the present work was high enough to obtain readings several folds greater than the reading of control samples (unstimulated). The sample blanks were also lowest for perfusates of stimulated and non-stimulated samples. The content of CA in the perfusate was expressed in terms of norepinephrine (base) equivalents.
Statistical analysis
The statistical significance between groups was determined by the Student's t- and ANOVA- tests. A p-value of less than 0.05 was considered to represent statistically significant changes unless specifically noted in the text. Values given in the text refer to means and the standard errors of the mean (S.E.M.). The statistical analysis of the experimental results was made by computer program described by Tallarida and Murray [36].
Drugs and their sources
The following drugs were used: acetylcholine chloride, norepinephrine bitartrate, nicardipine hydrochioride, amiloride hydrochloride hydrate, 3.4.5-trimethoxy benzoic acid 8-(diethylamino) octylester (TMB-8), pirenzepine hydrochloride, ethyleneglycol tetraacetic acid (EGTA), Ki16425, and Oleoyl-L-α-lysophosphatidic acid sodium salt from Sigma-Aldrich Chemical Co., U.S.A., and chlorisondamine chloride from Ciba Co., U.S.A. Gintonin, devoid of ginseng saponins, was prepared from Panax ginseng according to previously described methods [32], and supplied by professor Seung-Yeol Nah who is a professor of Konkuk University, Seoul, Korea. Gintonin was dissolved in deionized water and then diluted with Krebs solution before use. LPA and Ki16425 was dissolved in DMSO and then diluted with Krebs solution before use. Drugs were dissolved in distilled water (stock) and added to the normal Krebs solution as required. Concentrations of all drugs used are expressed in terms of molar base.
RESULTS
The secretory effect of CA evoked by gintonin
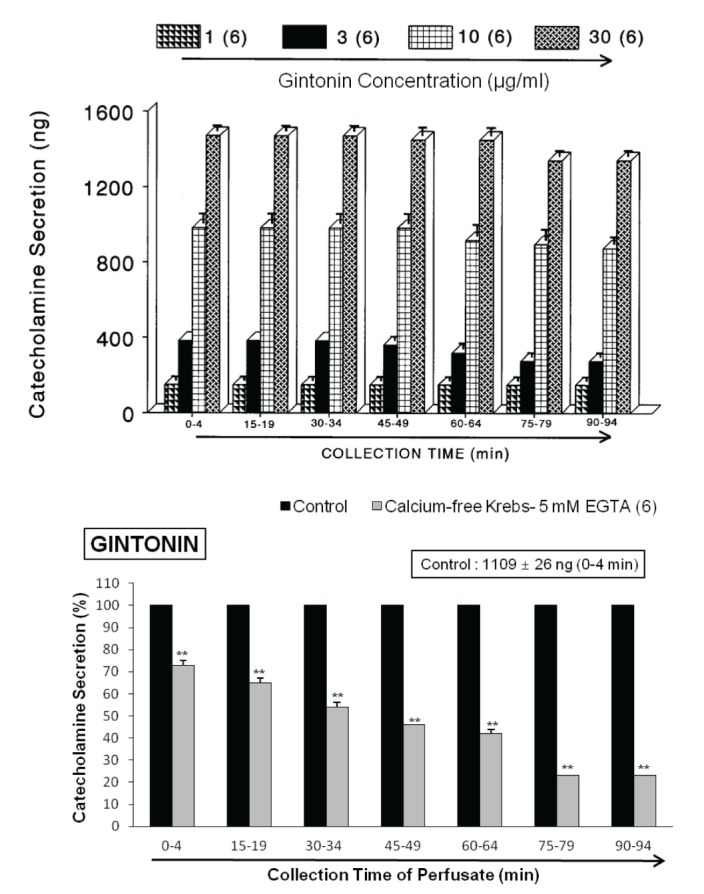
When the adrenal gland was perfused with oxygenated Krebs-bicarbonate solution for 60 min before experimental protocol is initiated, the spontaneous CA secretion reached steady state. The basal CA release from the perfused rat adrenal medulla amounted to 21±2 ng for 2 min from 10 experiments. The releasing effects to the perfusion of gintonin for 4 min are shown in Fig. 1 (upper). Infusion of gintonin (1~30 µg/mL) into the perfusion stream for 4 min at 15 min-intervals gradually dose-dependently exerted significant CA secretory responses from 149±21 ng to 1451±43 ng over the background release. These observations are largely consistent with results described previously that ginseng saponins cause the increased CA secretion from the isolated perfused rabbit adrenal glands [123] while rather opposite to the inhibitory effects on the CA secretion in rat adrenal gland [4] and SHRs [5], and in bovine adrenal chromaffin cells [67937].
In order to examine the tachyphylaxis to gintonin-evoked CA releasing effects, gintonin at the concentration of 10 µg/mL was given into the perfusion stream for 4 min seven times consecutively at 15 min-intervals, respectively. In 6 rat adrenal glands, there was no statistically significant difference in CA secretion (1344±29 ng~1451±43 ng) between periods (Fig. 1-upper). Therefore, in all subsequent experiments, 10 µg/mL of gintonin was used.
Effects of perfusion of Ca2+-free Krebs, nicardipine, TMB-8 and amiloride on gintonin-induced CA secretion
Since the physiological secretion of CA and dopamine-β-hydroxylase from the perfused cat adrenal gland is found to be dependent on the extracellular calcium concentration [38], it was of particular interest to test whether gintonin-induced CA secretory effect is also associated to extracellular calcium ions. Thus, the adrenal gland was perfused with calcium-free Krebs solution containing 5×10–3 M EGTA for 20 min prior to introduction of gintonin. Under the perfusion of calcium-free Krebs solution containing EGTA in 6 rat glands, gintonin (10 µg/mL)-evoked CA secretory responses were significantly inhibited to from 73±2% (0~4 min, p<0.01) to 23±1% (75~94 min, p<0.01) of the corresponding control response (1109±26 ng/0~4 min), as shown in Fig. 1 (lower).
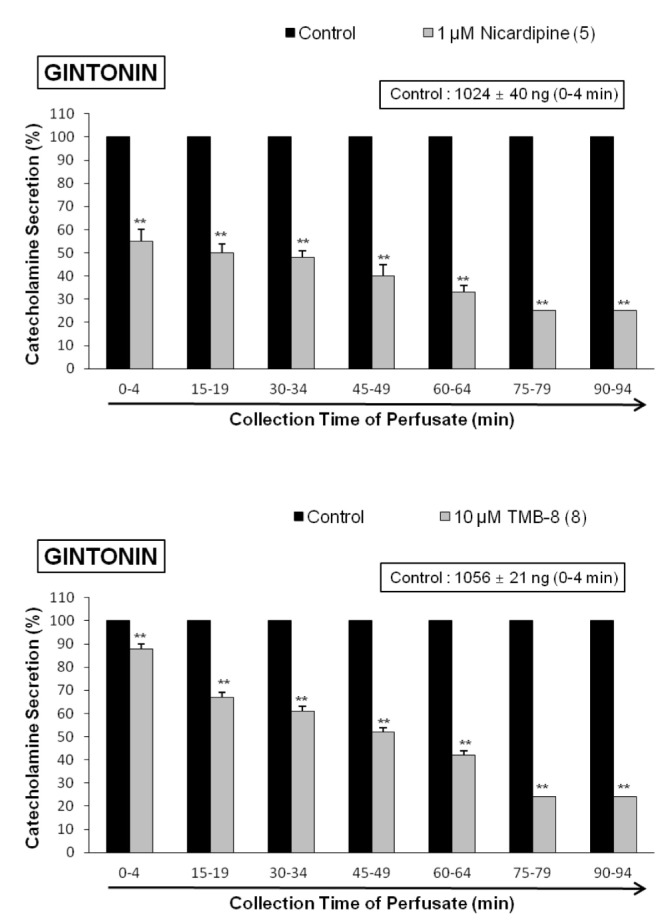
In order to investigate the effect of nicardipine, which is known to be a voltage-dependent L-type Ca2+ channel blocker and a dihydropyridine derivative [39], on gintonin-evoked CA secretion, nicardipine (10–6 M) was preloaded into the adrenal gland for 20 min. In the presence of nicardipine effect, gintonin (10 µg/mL)-induced CA was markedly reduced from 73±2% (0~4 min, p<0.01) to 23±1% (75~94 min, p<0.01), as compared with the corresponding control response (1024±40 ng/0~4 min) in 5 rat glands. Fig. 2 (upper) shows that nicardipine inhibits gintonininduced CA secretory responses relatively in time-dependent fashion.
It has been reported that muscarinic, but not nicotinic activation causes CA secretion independent of extracellular calcium in the perfused cat adrenal glands [40]. It suggests that the presence of an intracellular calcium pool is linked to muscarinic receptors, and that TMB-8, an intracellular calcium antagonist, inhibits both nicotinic and muscarinic stimulation-induced CA release in the rat adrenal glands [41]. Therefore, it was attempted to examine the effects of TMB-8 on gintonin-induced CA secretion. In 8 rat adrenal glands, gintonin (10 µg/mL)-induced CA secretion after preloading with TMB-8 (10 µM) for 20 min were greatly reduced from 88±2% (0~4 min, p<0.01) to 24±2% (75~94 min, p<0.01) in comparison with their corresponding control response (1056±21 ng/0~4 min), as shown in Fig. 2 (lower).
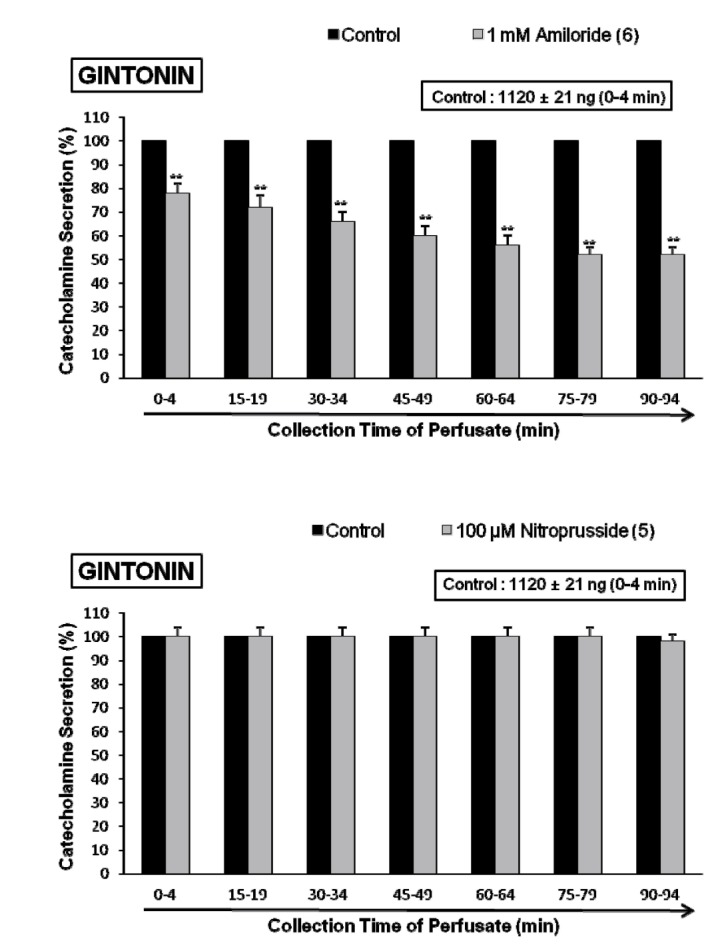
It was found that gintonin-evoked CA secretion is greatly inhibited by treatment with Ca2+-free Krebs, nicardipine or TMB-8, respectively. To examine the involvement of Na+ channel, Na+/Ca2+ exchanger and Na+/H+ exchanger in gintonin-induced increase in CA secretion, adrenal glands were treated with their specific inhibitor. Perfusion of adrenal gland with amiloride, which is known Na+/Ca2+ exchange inhibitors [4243], for 90 min caused a significant decrease in gintonin-induced CA secretion, as shown in Fig. 3 (upper). In the presence of amiloride (1 mM), gintonin (10 µg/mL)-induced CA secretion was markedly reduced from 78±4% (0~4 min, p<0.01) to 52±3% (75~94 min, p<0.01) relatively in time-dependent fashion, as compared with the corresponding control response (1120±21 ng/0~4 min) from 6 rat adrenal glands.
Effects of nitroprusside, chlorisondamine and pirenezepine on gintonin-induced CA secretion
In 5 rat adrenal glands, to study the relationship between NO and gintonin-induced CA secretion, sodium nitroprusside (100 µM) was perfused into the adrenal gland for 90 min. In the presence of sodium nitroprusside, gintonin (10 µg/mL)-induced CA secretory response was not affected as compared with its control secretion as shown in Fig. 3 (lower). There was no statistical significance in difference between the gintonin-evoked CA secretory responses before and after pretreatment with nitroprusside.
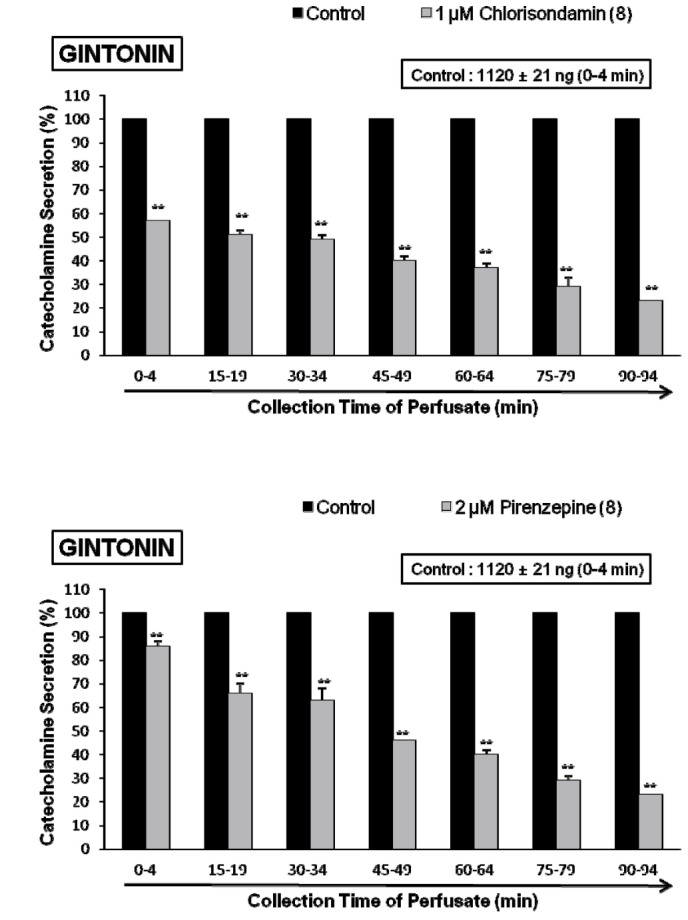
In order to certify the effect of chlorisondamine, a selective neuronal nicotinic receptor antagonist, on gintonin-induced CA release, the rat adrenal gland was loaded with chlorisondamine (1 µM) for 90min. In the presence of chlorisondamine, gintonin (10 µg/mL)-induced CA seretion were greatly inhibited from 57±2 ng (0~4 min) to 23±1 ng (90~94 min) of the control release (1120±20 ng/0~4 min) from 8 glands as shown in Fig. 4 (upper).
In order to observe the effect of muscarinic receptors, it would be interesting to examine the effect of pirenzepine on gintonin-induced CA secretion. In the present work, gintonin-induced CA output was significantly reduced in the rat adrenal gland preloaded with pirenzepine (2 µM) for 90 min. In 8 rat adrenal glands, gintonin (10 µg/mL)-induced CA secretory responses after treatment with pirenzepine were greatly inhibited to 86~23% of the control release (1120±20 ng/0~4 min) as shown in Fig. 4 (lower).
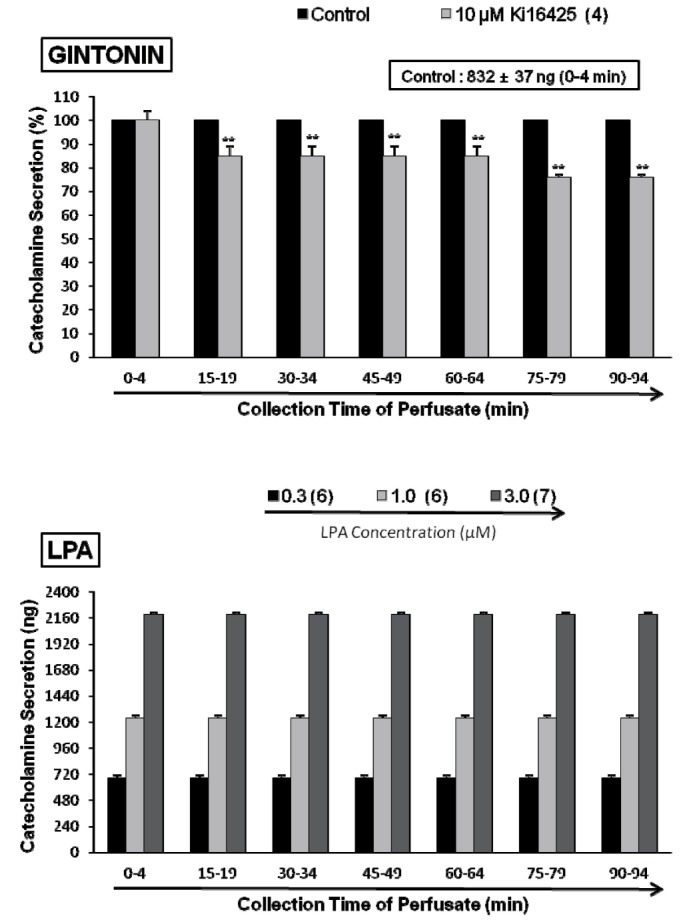
Effects of Ki16425 on lysophosphatidic acid (LPA)- and gintonin-induced CA secretion
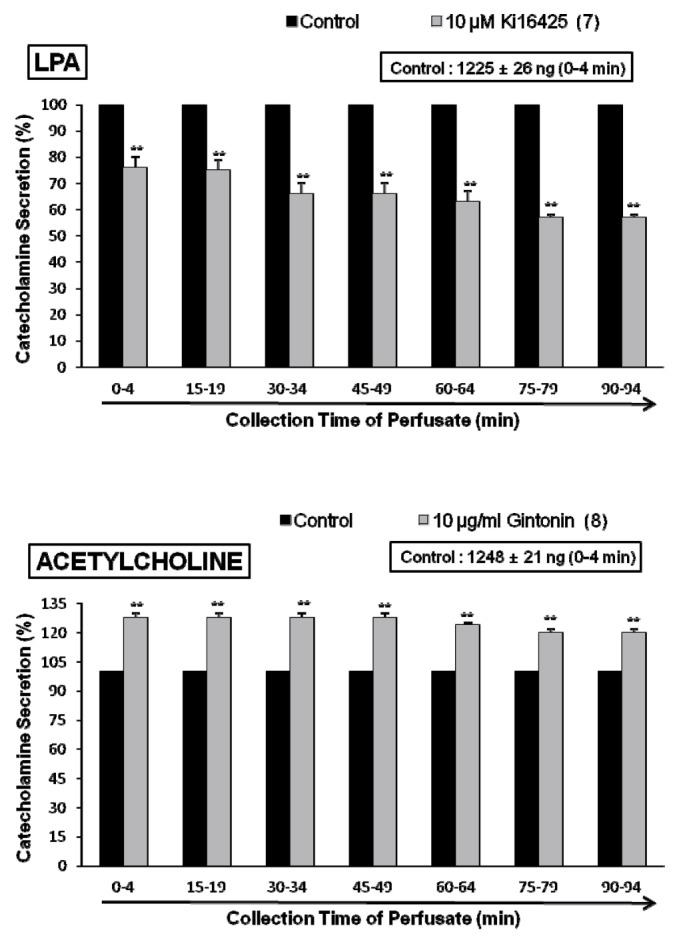
Since it has been previously reported that ginseng contains a novel G protein-coupled lysophosphatidic acid (LPA) receptor ligand, known as gintonin [303132], it is very interesting to examine the effect of LPA receptor antagonist on gintonin-induced CA secretory response. As shown in Fig. 5 (upper), in the presence of Ki16425 (10 µM), selective antagonist of LPA receptors [44], gintonin(10 µg/mL)-induced CA secretory responses were significantly inhibited to 85±4% (15~64 min, p<0.01) and 76±1% (75~94 min, p<0.01) as compared to their control secretion (832±37 ng/0~4 min). Moreover, in order to examine whether LPA can evoked CA secretion from the perfused art adrenal medulla, perfusion of LPA (0.3~3.0 µM) into the adrenal gland for 4 min at 15 min-intervals greatly enhanced the CA secretion from 683±27 ng (0.3 µM) to 2194±18 ng (3.0 µM) over the background release (21±2 ng) in a dose-dependent fashion (Fig. 5-lower). However, in the presence of Ki16425 (10 µM), LPA (1 µM)-induced CA secretory responses were markedly reduced to 85±4% (p<0.01)~57±4% (p<0.01) as compared to their control secretion (1225±26 ng/0~4 min) as shown in Fig. 6 (upper).
The effect of gintonin perfusion on ACh-induced CA release
In terms of the facts that gintonin-induced CA release was markedly inhibited by the pretreatment with chlorisondamine and pirenzepine as in Fig. 4, it was of interest to examine the influence of gintonin perfusion on ACh-induced CA secretory responses. The adrenal gland was perfused with gintonin (10 µg/mL) for 90 min. In the presence of gintonin, ACh-induced CA output was greatly enhanced to 128±2% (0~49 min, p<0.01), 124±1% (60~64 min, p<0.01) and 120±2% (75~94 min, p<0.01) as shown in Fig. 6 (lower).
DISCUSSION
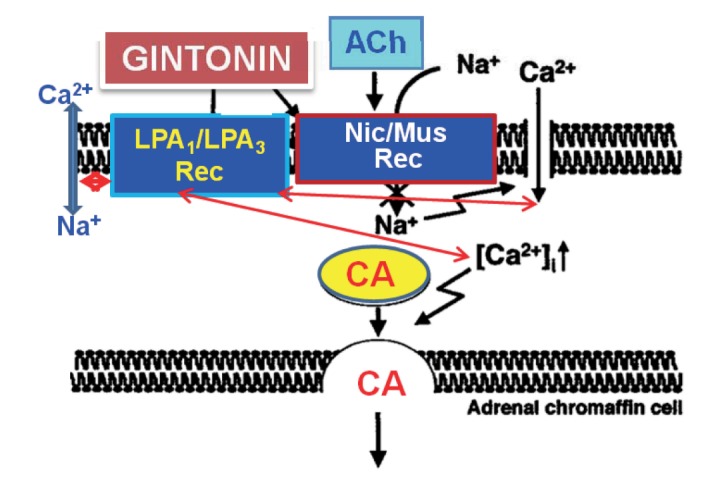
This is the first report demonstrating that gintonin greatly enhances CA secretion in a calcium-dependent fashion from the isolated perfused model of the rat adrenal medulla through activation of LPA (LPA1 and LPA3) receptors as well as neuronal cholinergic (nicotinic and muscarinic) receptors located on the rat adrenomedullary chromaffin cells.
In the present study, gintonin-evoked CA secretory responses were significantly inhibited in the presence of Ki16425, the LPA1/3 receptor antagonist. Moreover, LPA also greatly caused the facilitation of CA secretion in the perfused rat adrenal medulla in a dose-dependent fashion. However, this LPA-induced CA secretion was also markedly impeded in the presence of Ki16425. It is well-known that Ki16425 selectively inhibits LPA receptor-mediated actions, especially through LPA1 and LPA3; therefore, it may be useful in evaluating the role of LPA and its receptor subtypes involved in biological actions [44]. Therefore, these results indicate that gintonin-evoked CA secretion might be at least due to activation of LPA receptors located on adrenomedullary chromaffin cells.
In support of this idea, gintonin is found to be a novel G protein-coupled LPA receptor ligand [303132]. More recently, it has been shown that gintonin-mediated intracellular Ca2+ transients and dopamine release via LPA receptor activation in PC12 cells could be one mechanism of gintonin-mediated inter-neuronal modulation in the nervous system [33]. Furthermore, LPA is also known to facilitate dopamine release from PC12 cells, indicating that LPA stimulates dopamine release via specific LPA receptors presumably by activating the IP3 mediated Ca2+ release [45]. Also, Pan and his co-workers suggested the differential effects of LPA and S1P in releasing Ca2+ from the intracellular Ca2+ stores and modulating the stimulus-secretion coupling in bovine chromaffin cells [46]. In view of these findings, the present study demonstrates that gintonin can enhance the CA secretion from the perfused rat adrenal medulla through activation of LPA1/3 receptors.
In the present work, upon the repeated administration of gintonin at 15 min intervals into the perfused rat adrenal gland, the CA secretory response was no changed. Tachyphylaxis to releasing effects of CA evoked by gintonin was not observed by the repeated administration of concentrations used in this study.
Generally, the adrenal medulla has been employed as the model system to study numerous cellular functions involving not only noradrenergic nerve cells but also neurons. One of such functions is neurosecretion. During the neurogenic stimulation of the adrenal medulla, ACh is released from the splanchnic nerve endings and activated cholinergic receptors on the chromaffin cell membrane [47]. This activation triggers a series of events known as stimulus-secretion coupling, culminating in the exocytotic release of CA and their components of the secretory vesicles into the extracellular space. ACh, the physiological presynaptic transmitter at the adrenal medulla, releases CA and dopamine-β-hydroxylase by calcium-dependent secretory process [3848].
In the present work, the gintonin-induced release of CA seems to be due to exocytosis of CA storage vesicles subsequent through activation of nicotinic ACh receptors in the rat adrenomedullary chromaffin cells, because it was greatly abolished in the presence of chlorisondamine. Chlorisondamine is known to be a selective antagonist of neuronal nicotinic cholinergic receptors [39]. Also, in this study, the gintonin-evoked CA secretory response was significantly inhibited in the presence of pirenzepine, which is a selective antagonist of muscarinic M1-receptors. This finding indicates that gintonin-evoked CA release is mediated at least by stimulation of muscarinic ACh receptors. In general, subtypes of muscarinic receptors have been recognized in many tissues [49]. Receptor binding studies have supported the classification of muscarinic receptors into M1 and M2 on the basis of the selectivity profile of pirenzepine; receptors with a high affinity for pirenzepine are designated as M1 and those with low affinity as M2 receptors [5051]. Doods and his colleagues [52] have classified muscarinic receptors into M1 (pirenzepine sensitive, neuronal), M2 (cardiac) and M3 (smooth muscle and glandular). It has been shown that that combination of extracts of ginseng and ginkgo biloba significantly increased the level of ACh in whole brain of amyloid beta-protein treated rats [53]. This ginseng-induced improvement of behavioral impairment in amyloid beta-protein treated rats seems to be due to the increased ACh level, which results from the decreased AChE activities [53]. Su and his coworkers [54] have suggested that ginseng root has the ability to increase the release of ACh from nerve terminals in wild rats so as to stimulate muscarinic receptors activity located in the pancreatic cells for the secretion of insulin, which in turn lower plasma glucose. More recently, Kim and his coworkers [55] reported that activation of G protein-coupled LPA receptors by gintonin is coupled to the regulation of cholinergic functions. Furthermore, they also found that gintonin could be a novel agent for the restoration of cholinergic system damages due to amyloid beta-protein and could be utilized for prevention or therapy of Alzheimer's disease. In view of above studies, gintonin-evoked CA release seems to be mediated partly through activation of nicotinic as well as muscarinic receptors in the perfused rat adrenal medulla.
In the present study, gintonin-evoked CA releasing effect was not influenced in presence of sodium nitroprusside, known as an NO donor. This result demonstrates that gintonin-evoked CA secretion is not relevant to NO release in the perfused rat adrenal medulla.
The indispensable role of calcium in the neurosecretory process has been well established. Calcium is well-known to play the crucial role in the process of depolarization-neurotransmitter release coupling in many types of secretory cells [565758]. Furthermore, it has been found that nicotinic (but not muscarinic) stimulation also releases soluble ACh from the chromaffin cells by a calcium-dependent mechanism [59]. The activation of nicotinic receptors stimulates CA secretion by increasing Ca2+ entry through receptor-linked and/or voltage-dependent Ca2+ channels in the perfused rat adrenal glands [60] and isolated bovine adrenal chromaffin cells [616263].
In the present study, removal of extracellular Ca2+ markedly depressed gintonin-evoked CA secretion. Furthermore, treatment of nicardipine, a dihydropyridine derivative and L-type Ca2+ channel blocker, also blocked the gintonin-evoked CA release. The secretory effect of gintonin seems to be apparently dependent on extracellular Ca2+. However, in the present study, the reason for considerable response to gintonin in Ca2+ -free Krebs plus EGTA solution as well as in the presence of nicardipine is not clear. However, in the presence of TMB-8, an inhibitor of the intracellular Ca2+ store, gintonin-evoked CA secretion was greatly inhibited in this perfused rat adrenal medulla. This result indicates that gintonin-evoked CA secretion is associated with intracellular Ca2+ release from endoplasmic reticulum (Ca2+ store) of the rat adrenal chromaffin cells. In support of this notion, TMB-8 is also known to inhibit caffeine-induced 45Ca2+ release from, but not the uptake of 45Ca2+ in isolated bovine adrenomedullary cells [6465]. Moreover, it has been shown that caffeine-evoked CA secretion is also inhibited from the perfused adrenal gland of the cat [66] and the rat [67] in the absence of extracellular calcium. TMB-8 reduced the resting [Ca2+]i and attenuated histamine-, 5-HT-, and glutamate-induced increases of [Ca2+]i in basilar artery smooth [68] as well as in brain cells [69]. TMB-8 is reported to inhibit the Ca2+ release from intracellular stores or increased the up-take of Ca2+ into sarcoplasmic reticulum and the inhibition of Ca2+-influx from extracellular site may be an indirect mechanism [70]. Recently, it has also been shown that gintonin-mediated [Ca2+]i transients are coupled to the stimulation of dopamine release in PC12 cells [33]. Thus, in light of these findings, our result shows that gintonin can trigger Ca2+ release from the intracellular Ca2+ store of the rat adrenomedullary chromaffin cells that participates in the CA secretion. Such a store may not be easily depleted by mere removal of extracellular calcium or voltage-dependent L-type Ca2+ channel blocker.
In the present study, in the presence of gintonin (continuous infusion), ACh-evoked CA secretion were significantly potentiated. This result indicates that mechanism of gintonin-evoked CA secretion is quite different from that of Ach-evoked CA secretion.
In the present study, gintonin-evoked CA secretion was clearly shown to be inhibited markedly by treatment of arniloride in the perfused rat adrenal medulla. This result suggests that the CA secretion evoked by gintonin is probably due to the activation of Na+/Ca2+ exchange process, resulting from the inhibition of Na+/K+-ATPase in nerve endings. In support of this idea, it has already been reported that amiloride inhibits Na+/Ca2+ exchanger in various cells and tissues [717273]. In view of these findings, amiloride is considered to inhibit gintonin-evoked CA secretion, presumably through its blocking action on the Na+/Ca2+ exchange system, which may be stimulated by the elevation of intracellular Na+ concentration, resulting from the inhibition of Na+/K+-ATPase in the plasma membranes. Moreover, LPA has also been found to stimulate extracellular Na+-dependent 45Ca2+ efflux from cultured bovine adrenal chromaffin cells, probably through its stimulatory effect on membrane Na+/Ca2+ exchange [74]. In light of these findings, gintonin-evoked CA secretory response seems to be relevant to activation of Na+/Ca2+ exchange in the rat adrenal chromaffin cells.
However, in contrast to above the described results, Pan and his coworkers [75] have found that both sphingosine-1-phosphate and LPA inhibit inward Ca2+ current in a concentration-dependent manner in isolated bovine chromaffin cells. They also showed that LPLs reduce repetitive firing of action potentials (Aps) and inhibit both exocytosis and endocytosis, indicating that S1P and LPA at physiological concentrations attenuate the excitability of chromaffin cells and hint at the possible blockade of CA secretion induced by sympathetic neurons. Therefore, it seems that there is some difference between cultured bovine chromaffin cells and perfused rat adrenal medulla in exocytosis evoked LPA and gintonin. Further experiments will be pursued to characterize how gintonin and LPLs modulate the exocytosis, especially in the isolated perfused model of the rat adrenal medulla in near future.
Previously, it has been reported that some ginsenosides, including Rb2 and Rg3 as well as total ginseng saponin inhibit CA release evoked by stimulation of cholinergic receptors through inhibition of influx of Na+ and Ca2+ as well as reduction of intracellular Ca2+ release from its store, which is relevant to elevation of NO release [456789]. Whereas gintonin known as a novel G protein-coupled LPA receptor ligand [303132] enhances CA release through facilitation of influx of Na+ and Ca2+ as well as intracellular Ca2+ release via stimulation of cholinergic and LPA receptors. Thus, gintonin-mediated regulation of CA release is quite different from that of ginsenosides in the perfused rat adrenal medulla. Future studies will remained to be elucidated what the clinical roles of these two components are in their differential regulation of CA release.
As shown in Fig. 7, taken together, these results demonstrate the first evidence that gintonin increases the CA secretion from the perfused rat adrenal medulla in a dose-dependent fashion. This facilitatory effect of gintonin seems to be associated with activation of LPA- and cholinergic-receptors, which are relevant to the cytoplasmic Ca2+ increase by stimulation of the Na+ and Ca2+ influx as well as by the inhibition of Ca2+ uptake into the cytoplasmic Ca2+ stores, without the increased nitric oxide (NO). Based on these results, it is thought that gintonin, one of ginseng components, can elevate the CA secretion from adrenal medulla by regulating the Ca2+ mobilization for exocytosis, suggesting facilitation of cardiovascular effects. Also, these findings demonstrate that gintonin might be at least one of ginseng-induced hypertensive components.
 XML Download
XML Download