

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Neuroinflammation with microglial activation plays a role in neurodegenerative diseases, such as Alzheimer's disease and multiple sclerosis [1], and microglia involved in the inflammation of the central nervous system have neuroprotective effects [234]. Recent in vivo and in vitro studies have suggested that microglia are essential in neural development, synapse formation, and neuroinflammation involved in the impairment of adult neurogenesis [567]. This evidence was supported by the Gram-negative bacterial immune stimulation lipopolysaccharide (LPS)-induced neuroinflammatory model, which showed that LPS-induced activation of the Toll-like receptor 4 (TLR4) in microglia led to the production of the pro-inflammatory cytokines, IL-1β, IL-6, and TNF-α through the MAPK and NFκB pathways in BV2 cell lines [89]. In addition to the immune response by activated microglia, the neuroprotective cytokines, IL-4 and IL-25, are also produced [1011]. Moreover, other studies showed that adult neurogenesis in the hippocampus was affected through the TLR and NF-κB pathway in microglia [1213].
Adult neurogenesis is a process in which new neurons are continuously generated throughout adult life from the neural stem cells (NSCs) in the mammalian brain. Small populations of neurons continue to be born in the adult subventricular zone, the olfactory system, and the DG of the hippocampus. The neurogenic niches are important for the NSCs to have self-renewal and multipotent properties that can give rise to neurons, astrocytes, and oligodendrocytes [141516]. The neuronal fate specification (neurogenesis) and glial fate specification (gliogenesis) of the NSCs is determined by cellular communication within the microenvironment [17]. The newborn neurons are critical to cognitive functions including learning and memory. For instance, physical activity and an enriched environment, which are associated with improved memory function and synaptic plasticity, enhance adult neurogenesis in the DG [1819]. Furthermore, adult neurogenesis in the DG declines with aging and neurodegenerative diseases, which in turn causes cognitive deficits [202122].
Green tea has been shown to function as an anticancer reagent that inhibits the development and progression of skin, lung, mammary gland, and gastrointestinal tract tumors in animal models [2324]. Among the various polyphenols in green tea including epigallocatechin-3-gallate (EGCG), (–)-epigallocatechin, (–)-epicatechin, (+)-gallocatechin, and other catechins, EGCG is the most abundant and the most biologically active component [252627]. However, the recovery effects of EGCG on adult neurogenesis following neuroinflammation at the DG in the hippocampus are still unknown. In the present study, we investigate the effect of EGCG on impaired adult neurogenesis in a mouse model of LPS-induced neuroinflammation.
Here, we present the effects of EGCG on impaired adult hippocampal neurogenesis in vivo caused by neuroinflammation. We induced inf lammation by administering LPS in a mouse model and explored the positive effects of EGCG in NSC proliferation and neural differentiation using an immunohistochemical approach to detect bromodeoxyuridine (BrdU, an S-phase marker of proliferating cells), doublecortin (DCX, a marker of immature newborn neurons), neuronal nuclei (NeuN, a marker of mature neurons), ionized calcium binding adapter molecule 1 (Iba-1, a marker of microglia), and the levels of cytokines.
METHODS
Animals & chemicals
7-week-old male C57BL/6 mice were purchased from Daehan Biolink (Chungbuk, Korea). The animals were kept under standard conditions of 22℃ and a humidity of 55% with a 12 hour light-12 hour dark cycle, and allowed free access to food and water. Mice were divided into the following three groups: saline-treated group (sham control), saline-treated with lipopolysaccharide (LPS, E.coli, serotype 055:B5, St. Louis, MO, USA) intracerebroventricular (I.C.V.) injection group, and epigallocatechin-3-gallate (EGCG)-treated with LPS I.C.V. injection group (n=5 mice/each group). Supplementary Figure 1 demonstrated the schematic experimental design of time courses with LPS, EGCG and BrdU injection followed by sampling the brain for immunohistochemistry. All experiments were approved by the Animal Care and Use Committee of Chonnam National University. All chemicals used in the experiments were supplied by Sigma (St. Louis, MO, USA).
LPS-induced neuroinflammation in the brain
Experimental mice were anesthetized with zoletil (VIRBAC, Milperra, Australia) (25 mg/kg) and Rompun (Bayer HealthCare, Mississauga, Canada) (10 mg/kg). LPS for a single unilateral stereotaxic injection was prepared in phosphate-buffered saline (PBS, pH 7.4) at a final concentration of 1.0 mg/ml and was injected into the brain (0.6 mm caudal to the bregma, 1.5 mm to the right of the bregma, 2.0 mm ventral to the bregma). Mice were injected with vehicle (sterile saline) or LPS (3 µg in 3 µl sterile saline) following brain surgery at a dose of 1 µg/min.
EGCG treatment
EGCG (0.5 mg/kg) prepared in physiological saline (PBS containing 0.9% NaCl) was injected intraperitoneally (i.p.) for three times with 6 hours interval at 3 hours after recovered from anesthesia for LPS-injection.
BrdU injection
BrdU (Sigma, St. Louis, MO, USA) was dissolved in physiological saline and injected i.p. (50 mg/kg). To examine the proliferation of NSCs in the DG, BrdU was injected into the animals once, 3 hours prior to sacrifice. To study the survival rate of newborn cells generated from NSCs and the differentiation rate of NSCs, BrdU was injected once daily for 5 consecutive days and the mice were sacrificed on 28th day after the final BrdU injection.
Tissue preparation
Animals were perfused transcardially with 0.1 M PBS, followed by fresh cold 4% paraformaldehyde (PFA) in 0.1 M PBS. The brains were removed and fixed overnight in 4% PFA in 0.1 M PBS at 4℃, and then washed for 6 hours in PBS at 4℃. Sucrose-saturated brains were then embedded in freezing media (O.C.T compound, Leica Biosystems, Richmond, USA), frozen in chilled isopentane (–25℃), and stored at –80℃ until sectioning. Brains were cryocut coronally at a thickness of 40 mm using a Cryostat (Model CM3050; Leica Microsystems, Richmond, USA) and stored in cryoprotectant solution (25% ethylene glycol, 25% glycerol, and 0.05 M sodium phosphate buffer, Na-PB) at –20℃ until immunohistochemical (IHC) processing.
Fluorescent immunohistochemical staining (IHC)
Sections were washed in Na-PB and mounted on charged slide glasses for IHC. The sections immunostained for BrdU were pretreated with 2 N HCl for 30 minutes at 37℃ and neutralized with PBS before incubation with primary antibodies. Sections were incubated for 60 minutes with 5% normal horse serum in 0.4% Triton X-100 in PBS (PBST). The sections were incubated overnight at 4℃ with primary antibodies in the same buffer solution. Primary antibodies were used at the following concentrations: rat anti-BrdU (1:200; Abcam, Cambridge, UK), goat anti-doublecortin (DCX, 1:100; Cell Signaling, Danvers, MA), mouse anti-neuronal nuclear antigen (NeuN, 1:100; Chemicon, Temecula, CA), and rabbit anti-ionized calcium-binding adapter molecule 1 (Iba-1, 1:100; Wako Chemicals USA, Inc., Richmond, VA). Sections were washed three times with PBST for 10 minutes at room temperature and blocked in PBST containing 5% horse serum for 30 minutes. Sections were then incubated for 2 hours with secondary antibodies conjugated to FITC (Jackson Immuno-Research, West Grove, PA) or CY3 (Jackson Immuno-Research, West Grove, PA). The sections were washed three times with PBST, and stained with 10 mg/ml 4'6'-diamidino-2-phenylindole (DAPI) (Sigma-Aldrich, St. Louis, USA) for 30 minutes before mounted.
Quantification and image analysis
IHC images were acquired with a confocal microscope (Zeiss, Thornwood, NY) equipped with an argon/krypton laser (488 nm), two helium/neon lasers (543 and 633 nm), and a coherent laser (Santa Clara, CA), using the 20x objective lens. Images were analyzed with the Image J (Ver.1.4, NIH, USA). Cells were counted in a defined frame size of 20~50 with 35 µm×35 µm in every selected section. Colocalizations were investigated by performing z-stack acquisitions and three-dimensional reconstructions with the LSM software (Zeiss, Thornwood, NY). Adobe Photoshop (Version CS6, Adobe Systems, San Jose, CA) was used to adjust the contrast and brightness.
Western blot analysis
Brain was homogenized in RIPA buffer (150 mM NaCl, 1.0% IGEPAL® CA-630, 0.5% sodium deoxycholate and 0.1% SDS in 50 mM Tris, Sigma, St. Louis, MO, USA). Proteins were separated by SDS-PAGE and transferred to a nitrocellulose membrane. The membranes were incubated with primary anti-TLR4, anti-RelA, anti-pRelA, and anti-Actin antibodies (Cell Signaling, Danvers, MA or Abcam, Cambridge, UK). The expression levels of protein were visualized and analyzed with Fusion FX (Vilber Lourmat, Eberhardzell, Germany).
Cytokine measurements
The levels of IL-1β, IL-6, and TNF-α in the DG were measured using a commercial ELISA kit (R&D Systems, Minneapolis, MN, USA) as per the manufacturer's instructions, and using quantitative real-time PCR (Bio-Rad Lab. Inc., CA, USA). In detail, the total RNAs were prepared using Trizol Reagent (Gibco BRL) and reverse transcription was performed with a RT system containing Moloney Murine Leukemia Virus reverse transcriptase (Promega, Madison, WI) in accordance with the manufacturer's instructions. PCR was performed in a Palm-Cycler thermocycler (Corbett Life Science, Sydney, Australia) and the product was resolved in a 1.2% agarose gel. Real time amplification of cDNA was conducted in a Rotor-Gene 3000 System (Corbett Research, Morklake, Australia) using the SYBR Green PCR Master Mix Reagent Kit (Qiagen, Valencia, CA). The PCR conditions were as follows: incubation for 5 minutes at 95℃, followed by 30 cycles of denaturation for 15 seconds at 95℃, annealing for 15 seconds at 62℃ and extension for 15 seconds at 72℃. The primers were as follows: mouse IL-1β; 5'-AGG AGA ACC AAG CAA CGA CA-3' and 5'-CTT GGG ATC CAC ACT CTC CAG-3', mouse IL-6; 5'-GCC TTC TTG GGA CTG ATG CT-3' and 5'-GCC TTC TTG GGA CTG ATG CT-3', mouse TNF-α; 5'-ATG GCC TCC CTC TCA TCA GT-3' and 5'-CTT GGT GGT TTG CTA CGA CG-3' and β-actin; 5'-GAT CTG GCA CCA CAC CTT CT-3' and 5'-GGG GTG TTG AAG GTC TCA AA-3'. The relative levels of mRNA were calculated using the standard curve generated from the cDNA dilutions. The mean cycle threshold (Ct) values from quadruplicate measurements were employed in the calculation of gene expression, with normalization to β-actin as an internal control. Calculation of the relative levels of gene expression was performed using Corbett Robotics Rotor-Gene software (Rotor-Gene 6 version 6.1, Build 90).
Data analysis
A complete series of 1 in 10 sections of the DG was analyzed. For each experiment, 9 sections per mouse were selected for analysis. The immunolabeled cells were counted and multiplied by 10 to obtain the total number of labeled cells throughout the DG. The data are expressed as the mean±SEM. Statistical analyses were performed using a one-way analysis of variance (ANOVA) followed by a Student-Newman-Keuls test for multiple comparisons. Results were considered statistically significant when the p value was less than 0.05.
RESULTS
EGCG recovered the proliferation of adult NSCs in the LPS-injured hippocampal DG
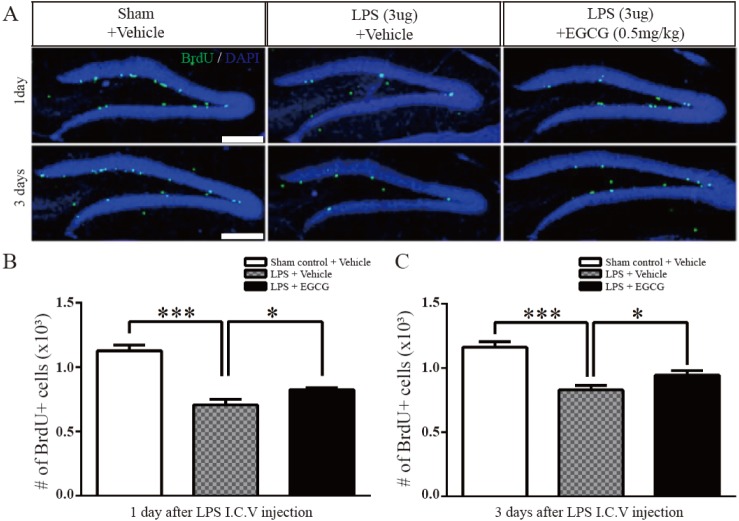
To study the effect of EGCG in the DG related to neurogenesis, the proliferation of adult NSCs in the DG was initially examined in an LPS-induced neuroinf lammation mouse model by administering LPS (3 µg/animal with 1 mg/ml stock) via intracerebroventricular (I.C.V.) injection (n=5 each). The number of BrdU (S-phase marker)-positive cells were compared at 3 hours following a single BrdU injection (i.p. 50 mg/kg), which indicates proliferating cells, and were found to be significantly decreased at 1 day (0.7066±0.04408×103, 38% of decreased compared to sham control, F (4.4)=1.076, p=0.0002, two-tailed unpaired one-way ANOVA) and day 3 (0.8292±0.03464×103, 29% of decreased compared to sham control, F (4.4)=1.577, p=0.0003, two-tailed unpaired one-way ANOVA) post-LPS injection in mice treated with either vehicle or EGCG compared with the sham controls. However, EGCG significantly improved the number of BrdU-positive cells affected by LPS in the hippocampal DG (day1; 0.8242±0.01520×103, 16% of increased compared with LPS only, F (4.4)=8.406, p=0.0357, day 3; 0.9462±0.03309×103, 14% of increased compared with LPS only, F (4.4)=1.096, p=0.0404, two-tailed unpaired one-way ANOVA) (Fig. 1). These results show that treatment with EGCG recovered NSC proliferation, which was impaired by LPS-induced neuroinflammation.
EGCG ameliorated the immature neuronal differentiation of adult NSCs in the LPS-injured hippocampal DG
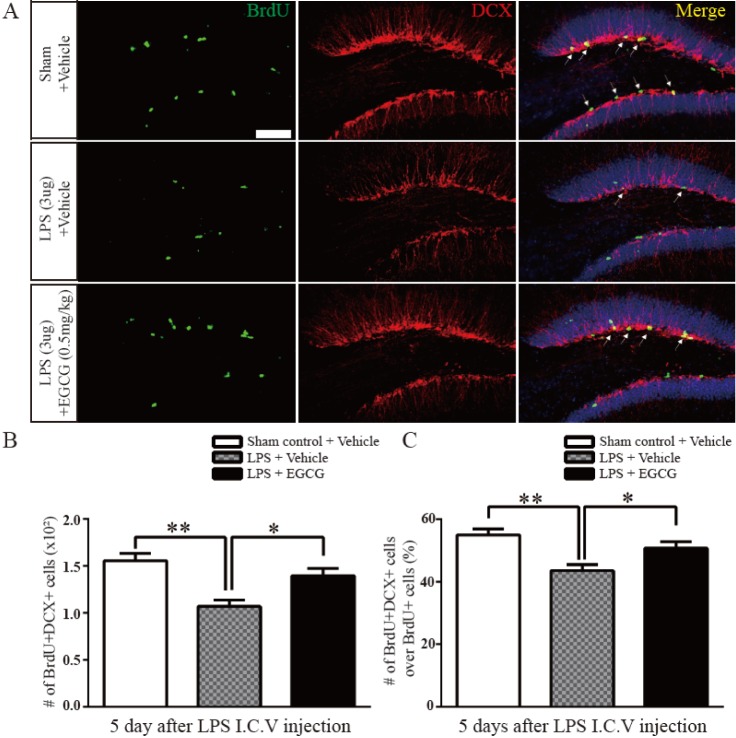
DCX (immature neuronal marker) is transiently expressed in the soma and dendrites of immature newborn neurons. The newborn NSCs are differentiated into neurons to serve as functional units in the nervous system following their proliferation. Thus, we studied whether EGCG ameliorated the differentiation of adult NSCs after LPS-induced injury by counting the number of BrdU- and DCX double-positive cells and the ratio of BrdU+ DCX+ cells/BrdU+ cells in the hippocampal DG 5 days after injection with BrdU (n=5 each). The number of double positive cells was decreased post LPS injection compared to that of sham controls (1.070±0.06641×103, 32% of decreased compared to sham control, F (4.4)=1.287, p=0.0013), but it was significantly improved after injection with EGCG in the LPS-injured group compared with the LPS-injured group with no treatment (1.398±0.07632×103, 30% of increased compared to LPS only, F (4.4)=1.321, p=0.0118) (Fig. 2A and B). Similarly, the ratio of the total number of BrdU- and DCX-double positive cells to the total number of BrdU-positive cells was also decreased in the LPS-treated group compared with the sham controls (43.60±1.887×103, 11.4% of decreased compared to sham control, F (4.4)=1.017, p=0.0026), but the ratio was recovered in the EGCG-treated LPS-injured group compared with the LPS-injured group with no treatment (50.80±1.985×103, 7.2% of increased compared to LPS only, F (4.4)=1.107, p=0.0302, two-tailed unpaired one-way ANOVA) (Fig. 2A and C). These results suggest that EGCG had a recovery effect on the neuronal differentiation of adult NSCs, which was impaired by LPS in the early stage.
EGCG amended the mature neuronal differentiation of adult NSCs in the LPS-injured hippocampal DG
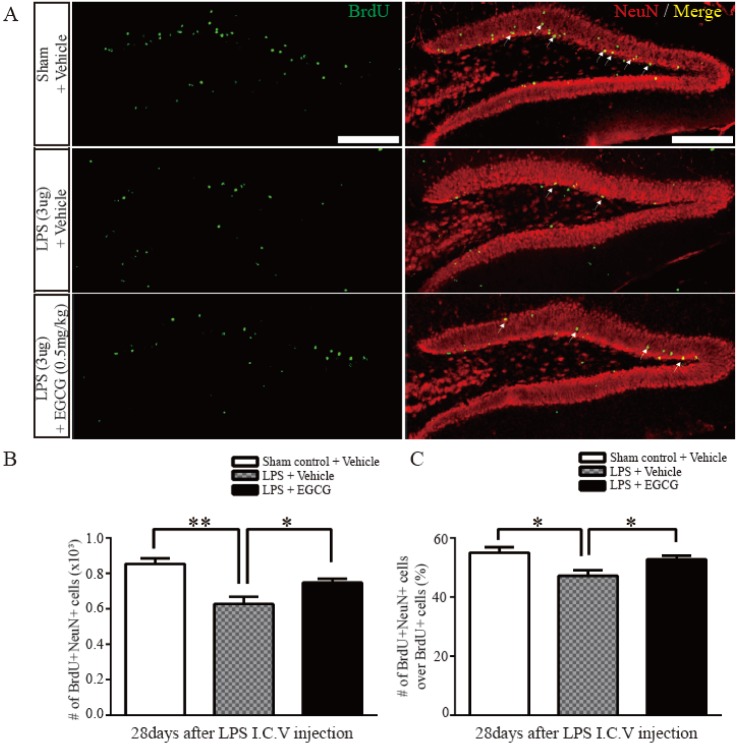
To investigate the effect of EGCG on the maturation of newborn neurons following LPS-induced neuroinflammation, immunohistochemical analysis was performed using antibodies against the mature neuronal marker, NeuN, and BrdU. The results show that the number of cells co-localized with NeuN and BrdU are decreased in the LPS-injured group (0.6280±0.03967×103, 27% of decreased compared to sham control, F (4.4)=1.623, p=0.0021), but the number of these cells are increased in the EGCG-treated LPS-injured group compared with the vehicle-treated LPS-injured group (0.7476±0.02307×103, 19% of increased compared to LPS only, F (4.4)=2.958, p=0.0313) (Fig. 3A and B). Furthermore, the ratio of NeuN/BrdU double-positive cells to the total BrdU-positive cells in the EGCG-treated LPS-injured group was significantly increased compared with that of the vehicle-treated LPS-injured group (52.80±1.158, 5.6% of increased compared to LPS only, F(4.4)=2.791, p=0.0378, two-tailed unpaired one-way ANOVA), which was decreased compared with the sham control group (Fig. 3C). These results support the notion of a beneficial effect of EGCG on mature neuronal differentiation of adult NSCs, which was impaired by LPS-induced neuroinflammation.
EGCG improved adult NSC survival in the LPS-injured hippocampal DG
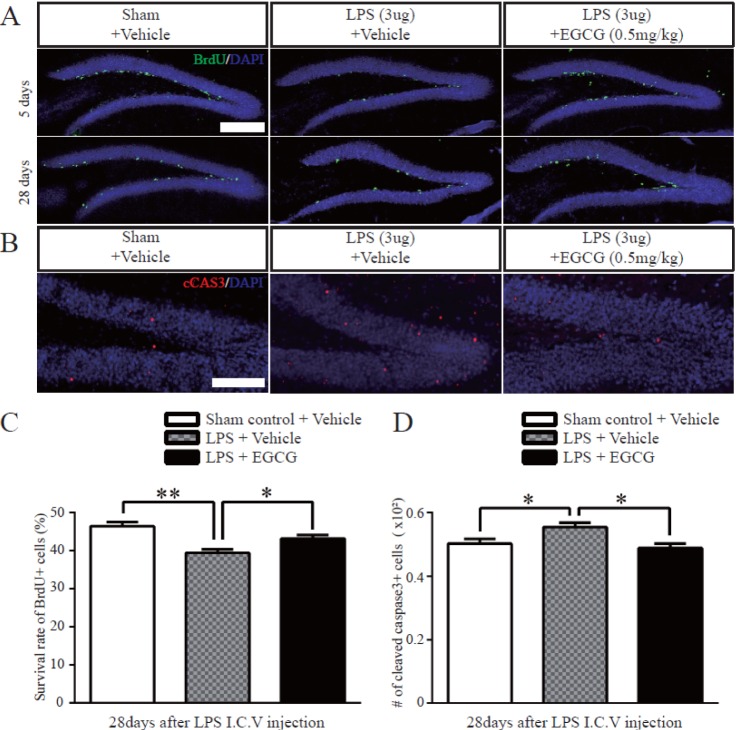
Following NSC proliferation in the DG, the newborn NSCs survive and differentiate into neurons. To determine the survival rate of newborn cells derived from NSCs in the DG, the number of BrdU-positive cells was quantified 3 hours and 28 days after the last injection of BrdU, which was injected once a day for 5 consecutive days. The total number of BrdU-positive cells was decreased in the LPS-injured group, and was recovered in the EGCG-treated LPS-injured group up to the level of the sham controls (n=5 each). Subsequently, we quantified the survival rate of newborn cells by counting the total BrdU-positive cells, 28 days after the final of 5 consecutive days of BrdU injections, to BrdU-positive cells, 3 hours after the final of 5 consecutive days of BrdU injections. The rate was decreased in all groups, but the EGCG-treated LPS-injured group showed an improved survival rate of newborn cells compared with the LPS-injured group with no treatment (43.20±0.8602, 3.8% of increased compared to LPS only, F (4.4)=1.162, p=0.0170, two-tailed unpaired one-way ANOVA) (Fig. 4A and C). Further, to examine the effect of EGCG on newborn cell survival related to apoptosis, the number of cleaved caspase 3-positive cells was counted. The total number of cleaved caspase 3-positive cells was significantly decreased in the EGCG-treated LPS-injured group compared with the vehicle-treated LPS-injured group (0.4880±0.01393×102, 12% of decreased compared to LPS only, F (4.4)=1.062, p=0.0109, two-tailed unpaired one-way ANOVA) (Fig. 4B and D). The results show that neuroinflammation decreased the survival of dividing cells, whereas EGCG recovered the survival of dividing cells, which was impaired by LPS-induced neuroinflammation.
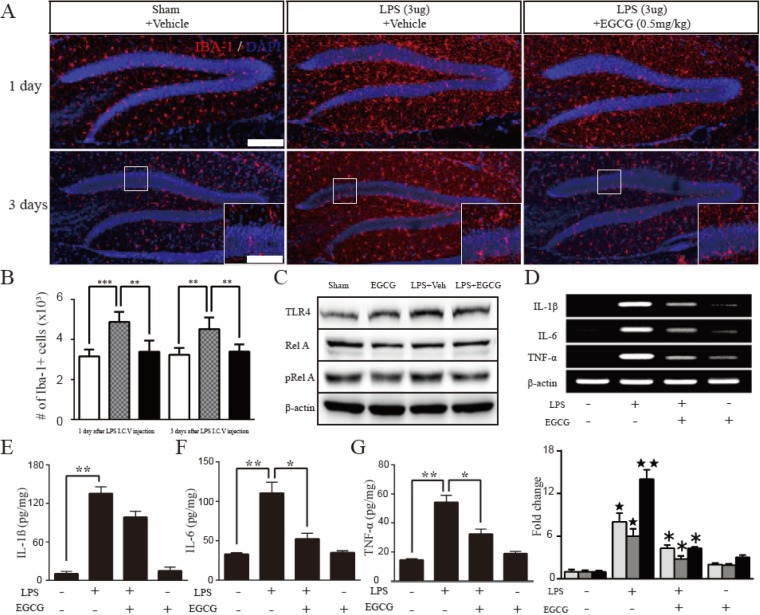
EGCG downregulated the expression of proinflammatory cytokines through the TLR4/NF-κB pathway in LPS-induced neuroinflammation
To study the effect of EGCG on neuroinflammation in detail, the activity of microglia following LPS I.C.V. injection was quantified by the immunoreactivity of Iba-1, a specific marker of microglia (n=5 each). The number of Iba-1-positive cells in the DG was increased (day 1; 4.876±0.2238×103, 54% of increased compared to sham control, F (4.4)=2.181, p=0.0002, day 3; 4.528±0.2567×103, 39% of increased compared to sham control, F (4.4)=2.917, p=0.0025, two-tailed unpaired one-way ANOVA ) by LPS injection, which was suppressed at day 1 and day 3 of EGCG treatment (day 1; 3.393±0.2497×103, 7% of decreased compared to LPS only, F (4.4)=1.245, p=0.0022, day 3; 3.402±0.1609×103, 4% of decreased compared to LPS only, F (4.4)=2.544, p=0.0059, two-tailed unpaired one-way ANOVA ) (Fig. 5A and B). Since LPS acted as a TLR4 agonist, the expression of TLR4, NF-κB, and cytokines was examined as molecular targets of EGCG in adult neurogenesis impaired by LPS-induced neuroinflammation. The expression of TLR4 and phospho-RelA (activated form) was upregulated in the LPS-injured group compared with the sham controls, but the expression of TLR4 and phospho-RelA was suppressed in the EGCG-treated LPS-injured group compared with the LPS-injured group. Therefore, LPS activated the expression of TLR4 and stimulated NF-κB, but EGCG attenuated the LPS-induced TLR4 and NF-κB (Fig. 5C). In addition, LPS upregulated the production of IL-1β, IL-6, and TNF-α by microglia, however, the increased mRNA and protein levels of cytokines induced by LPS injection were compromised by EGCG up to 53.75%, 46.67%, and 30.71% for IL-1β, IL-6 and TNF-α transcripts, respectively, and up to 72.61%, 47.59% and 59.57% for proteins, respectively, compared with the expression levels of the LPS-injured group (p<0.05) (Fig. 5D~G). The results demonstrate that EGCG suppressed the inflammatory activities of microglia, resulting in the attenuation of proinflammatory cytokine production via the TLR4/NF-κB signaling pathway.
DISCUSSION
Adult NSCs in the hippocampal DG have the ability to generate self-renewing neural stem cells possessing multipotency [28]. Adult neurogenesis occurs in three discrete stages, proliferation of NSCs, cell survival, and neuronal differentiation, which are affected by various factors such as hormones, growth factors, enriched environment, stress, drugs, and pathological stimulation [2930]. The process in the DG affected by these factors is associated with cognitive functions including learning and memory [31]. Neuroinflammation in the brain following injuries causes neural development-related diseases, which modulate adult hippocampal neurogenesis [3233]. During the inflammatory response following injury to the central nervous system (CNS), microglial cells and a population of glial cells of the CNS as immune cells produce proinflammatory cytokines. In particular, IL-6 and NO affect adult hippocampal neurogenesis, which are involved in the pathogenesis of neurological diseases such as Alzheimer's disease (AD) and Parkinson's disease (PD) [3435].
EGCG, among the catechins, a major subgroup of polyphenolic flavonoids in green tea, is well-known for its anti-carcinogenic effects via the suppression of the proliferation and angiogenesis of cancer cells [36]. Even though the instability of catechins, previous studies were showing that EGCG was effectively delivered over blood brain barrier and was founded physiologically activated forms in the brain [3738]. In addition, it may be used for the prevention and treatment of AD and PD based on the evidence that EGCG promotes adult hippocampal neurogenesis [394041]. Moreover, recent studies have demonstrated that EGCG had a neuroprotective effect through the reduction of neuroinflammation [42434445]. Oxidative stress has been implicated in the pathophysiology of the majority of neurodegenerative diseases, and most of the researches in this field have focused on EGCG as a natural pharmacological compound [46]. However, there have been no studies assessing the effect of EGCG on neurogenesis in the DG of the adult hippocampus impaired by neuroinflammation. Therefore, here, we investigated whether EGCG improves adult neurogenesis via the modulation of neuroinflammation in vivo.
In the present study, the effect of EGCG on the proliferation of adult NSCs in the DG following LPS injection was investigated. Proliferation of adult NSCs in the DG was inhibited by LPS injection, however, the number of BrdU-positive cells indicating the rate of adult NSC proliferation was improved within one day in the EGCG-treated LPS-injured group (Fig. 1A~C). Moreover, the survival rate of newborn cells, represented by the ratio of the total number of BrdU-positive cells 28 days after the final BrdU injection following LPS injection, was amended by EGCG treatment (Fig. 2A~C). These results suggest that EGCG rescued the proliferation and survival of adult NSCs in the DG impaired by LPS-induced neuroinflammation, which is consistent with that of previous reports showing the effective role of EGCG in increasing cell proliferation in mouse hippocampal DG and neural progenitor cell proliferation during adult hippocampal neurogenesis [4748].
To investigate the effect of EGCG on immature neuronal differentiation in the DG following LPS-induced neuroinflammation, immunohistochemical analysis was performed using a marker of immature neural differentiation, DCX, and BrdU, 5 days after BrdU injection following LPS with or without EGCG injection. The number of double positive cells (DCX+ and BrdU+) was decreased post LPS injection compared with that of the sham controls, however, it was significantly improved in the EGCGtreated LPS-injured group compared with the LPS-injured group with no treatment (Fig. 3A~C). The results indicate that EGCG ameliorated the neuronal differentiation of NSCs impaired by LPS in the adult hippocampal DG at an early stage. In addition, NeuN (a mature neuronal marker) and BrdU-positive cells at 28 days were decreased in the LPS-injured group, whereas the number of double positive cells was increased in the EGCG-treated LPS-injured group compared with the vehicle-treated LPS-injured group (Fig. 4A~C). It is indicating that EGCG had a recovery effect on the neuronal differentiation of adult NSCs at a late stage, which was impaired by LPS induced neuroinflammation. These results demonstrate that the neuronal differentiation of adult NSCs in the DG is rescued by the injection of EGCG into mice with LPS-induced neuroinflammation. Our data also was supported the beneficial effect of EGCG on adult neurogenesis by showing improved proliferation and differentiation of adult mouse hippocampal NSCs in the DG under normal conditions [48].
TLRs are expressed in immune and non-immune cells and are crucial for innate immune responses. For instance, TLR4 is activated by LPS-mediated inflammation in immune cells [49], and is also found in the central nervous system for regulating neurogenesis [50]. A previous report showed that EGCG has anti-inflammatory activity via the downregulation of TLR4 expression [51]. Thus, the recruitment of activated microglia was evaluated in LPS-induced neuroinflammation by showing the number of Iba-1-positive microglial cells. LPS activated the concentrating microglia in the brain, however, the activated microglia were suppressed by EGCG treatment, which modified the TLR4-mediated NF-κB pathway and proinflammatory cytokines (Fig. 5A~G). The results suggest a potential role of EGCG in significantly improving adult neurogenesis impaired by neuroinflammation, and show that the beneficial effect was associated with the downregulation of proinflammatory cytokines through the modulation of microglial activity.
It is still unclear whether EGCG decreases the level of oxidative stress in neuroinflammation for its improvement of adult neurogenesis. Published reports have demonstrated that microglia produce reactive oxygen species (ROS) in PD [5253], and in vivo and in vitro models of AD have shown that EGCG modulates the cellular mechanisms of neuroprotection and neurorestoration through the activation of the protein kinase C pathway for improving cell survival and apoptosis and antioxidant function against ROS [5455]. These works support that EGCG may be involved in the regulation of ROS for rescuing adult neurogenesis affected by neuroinflammation. To effectively address these questions, studies regarding the molecular mechanisms of EGCG in relation to ROS using primary cultures of adult NSCs are required in the future.
In conclusion, we suggest that the administration of EGCG was beneficial for improving the proliferation, survival rate, and neuronal differentiation of adult NSCs in the DG by suppressing the activity of microglia and the TLR4-mediated NF-κB pathway in impaired neurogenesis caused by neuroinflammation. Therefore, EGCG may be a potential therapeutic agent for neuroinflammatory diseases.
 XML Download
XML Download