

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Recently, the functionalized bio-active textile products have been increasingly developed [1,2,3]. For examples, the use of special silk clothes may be useful in the management of atopic dermatitis in children [4]. Lee et al. [5] showed that multi-functional fabric wear is effective in inducing deep sleep, increasing growth hormone, and improving the quality of sleep. Also, they suggested that multi-functional fabric may act to stabilize both the autonomic nervous system and psychological state [6]. GERMACOLOR (Germany) and VENTE (Korea) collaborated to develop the fabric radiating bio-active energy. The bio-active materials coated fabric (BMCF) was coated with bio-active materials containing over 30 kinds of minerals. The clothing made of this fabric has layer of bio-active energy which reacts with far infrared rays from human body and it is transferred into human body. Advanced research has shown the improvement characteristics in muscular maximal exercise performance and recovery from muscle fatigue when using this testing product. And, any kind of unusual skin reaction has not been detected in safety evaluation (data not shown). However, the study for intra-cellular mechanisms of signaling pathway induced by this functionalized textile has not been enough.
Aged muscles retain dysfunctional fiber segments which atrophy and break contributing to the loss of muscle mass and function [7]. Recent studies have shown that mitochondria play a major role in the regulation of myogenesis [8,9,10]. The abundance, morphology, and functional properties of mitochondria become altered when the myoblasts differentiate into myotubes [11,12,13]. Mitochondria are vital organelles for the energy production and for the regulation of signaling cascades in skeletal muscle. Mitochondria generate adenosine triphosphate (ATP), which is used as a source of chemical energy, and control the cell cycle and growth by involving in cellular signaling pathway [14]. Thus, a variety of disorders can be caused by oxidative stress, damage and subsequent dysfunction in mitochondria [15].
Apoptosis is mainly divided into two of pathway, extrinsic and intrinsic. The extrinsic pathway is occurred by cell surface death receptors such as Fas receptor (FasR). On the other hand, the intrinsic pathway is occurred by mitochondria-mediated signaling cascade, which release cytochrome c (cyt c) caused by mitochondrial permeability transition pore (mPTP) into cytosol [16,17]. Autophagy has crucial roles in the cellular adaptation to stress as a quality control mechanism [18]. Thus, autophagy has relation to an essential cytoprotective pathway and a potential anti-aging mechanism [19]. Autophagy may occur either as a general phenomenon, for instance when cells lack nutrients and mobilize their energy reserves, or it can specifically target cellular structures such as damaged mitochondria (mitophagy). Because of the danger of having damaged mitochondria in the cell, the timely elimination of damaged and aged mitochondria is essential for maintaining the integrity of the cell. This turnover process consists of the sequestration and hydrolytic degradation by the lysosome, a process also known as mitophagy. Mitochondrial biogenesis is characterized as a vital process in the synthesis and degradation of the organelle [20]. This fundamental process comprehends the synthesis import and incorporation of lipids and proteins to the existing mitochondrial reticulum; the stoichiometric assembly of multi-subunit protein complexes into a functional respiratory chain; replication of the mitochondrial DNA; selective degradation of mitochondria by autophagy [21,22,23].
In this study, we examined the effects of the BMCF on skeletal muscular mitochondria. To observe the effects, the fabric was put on old-aged rat then the oxygen consumption efficiency and copy numbers of mitochondria, and mRNA expression of apoptosis- and mitophagy-related genes were verified as times passed.
METHODS
Animals and groups
Sprague-Dawley rats (12 weeks, male) weighing 350±15 g were obtained from Samtako (Osan, Korea). The animals were allowed to acclimatize for 1 week before the experiments and were housed in a standard, controlled environment with a temperature of 22±3℃, humidity of 50±10%, 12 h light-dark cycle, and a ventilation 10~15 times/h with wind velocity of 10~20 cm/sec. The animals were fed filtered tap water and purified pellet diet (Dong A One Corporation Ltd., Korea). The protocols used conformed to the National Institutes of Health Guide for the Care and Use of Laboratory Animals and were approved by the Ethics Committee for Laboratory Animals of Chung-Ang University (approval number: 14-0023). The animals were divided into one control group (eight animals) and two experimental groups (eight animals each). Cloth made from conventional fabric (V-Con) was used for the control group while 10% (V-10) and 30% (V-30) BMCF were used for each experimental group.
BMCF
The fabric samples were provided by VENTEX. The bio-active material contained 30 kinds of minerals, such as SiO2, Mg, Al2O3, Na, Ca, and Fe2O3, and was blended with acrylic resin at the desired ratio and coated on one side of the fabric. GERMACOLOR and VENTEX collaborated to develop this fabric, and the clothing made from this fabric has a layer that radiates bio-active energy. The bio-active materials radiate unique energy generated according to the molecular structure and vibration of each element. The energy reacts with far infrared rays emanating from the human body and is transferred into the human body.
Mitochondria isolation
The animals were sacrificed through ether inhalation anesthesia and cardiac puncture. After euthanasia was administered, the subtrapezial muscle was rapidly removed and was washed at least three times in PBS. The mitochondria fraction was obtained according to the modified methods described by Frezza et al. [24]. The muscles were homogenized in a homogenizer (WiseTis, Korea) with 3 ml MIB1 (KCl 180 mM, Na2-EDTA 0.5 mM, Tris 10 mM, pH 7.4). The resulting homogenate was centrifuged at 1,000×g for 10 min at 4℃, and the supernatant was collected. The supernatant was centrifuged once again at 1,000×g for 10 min at 4℃, and the supernatant was again collected. Finally, the supernatant was centrifuged at 10,000×g for 10 min at 4℃, and the supernatant was discarded. A mitochondria-enriched pellet was re-suspended in 1 ml MIB2 (KCl 180 mM, Na2EDTA 0.5 mM, Tris 10 mM, BSA 1 g/L, pH 7.4) and was centrifuged at 10,000×g for 10 min at 4℃. The mitochondria-enriched pellet was then ready for the oxygen consumption measurement.
Oxygen consumption measurement
The mitochondrial oxygen consumption was measured according to the modified methods described by Frezza et al. [24]. The measurement of the respiratory control rates was performed with 20 mM ADP, G/M (5 mM Glutamate, 2.5 mM Malate), MIR05 (EGTA 0.5 mM, MgCl2-6H2O 3 mM, K-lactobionate 60 mM, taurine 20 mM, KH2PO4 10 mM, HEPES 20 mM, sucrose 110 mM, BSA 1 g/L, pH 7.1). Oxygen consumption was performed in a NeoFox chamber (Ocean Optics Inc., Dunedin, USA) and was monitored with a NeoFox viewer (version 2.30, Ocean Optics Inc.) program. The respiratory control ratio (RCR) is the ratio of respiration between the presence of ADP (state 3) and the absence of ADP (state 4).
Gene extraction and cDNA synthesis
DNA was extracted from rat subtrapezial muscle using the QIAamp DNA Mini Kit (Qiagen, Germany) according to the product instruction manual. The extracted DNA contained two kinds of DNA: genomic DNA (gDNA) and mitochondrial DNA (mtDNA). RNA was extracted from rat subtrapezial muscle tissue using TRIzol reagent (Invitrogen Life Technologies, USA) according to the product instruction manual. The quantity of total RNA was determined using a NanoDrop 1000 (Thermo Scientific, USA). The total RNA was reverse-transcribed to cDNA using the RNA to cDNA Ecodry™ premix (oligo dT) kit (TaKaRa, Japan) with 2,000 ng of RNA according to the product instruction manual.
Mitochondrial DNA copy number measurement
The sequences for the mitochondrial genome and Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) primers are shown in Table 1. Each real-time PCR reaction mixture included 0.4 µl MgCl2 (final concentration 2 mM), 0.5 µl forward primer (final concentration 0.5 µM), 0.5 µl reverse primer (final concentration 0.5 µM), 1 µl LightCycler DNA Master SYBR Green I 10×(Roche, Germany), DNA template (15 ng) and up to 10 µl of distilled water (DW). Real-time PCR was performed in a LightCycler 2.0 (Roche). The real-time PCR conditions were as follows: denaturation (95℃ for 10 min), a three-segment amplification program repeated 35 times (95℃ for 10 sec; 62℃ for 10 sec; 72℃ for 10 sec), melting curve program (65℃ to 95℃ with a heating rate of 0.1℃/sec) and cooling down (40℃). The results were expressed as the ratio between mtDNA and gDNA. The mtDNA and gDNA Ct ratios were compared according to the modified methods described by Guo et al. [25].
mRNA expression
The mRNA expression of apoptosis-related genes was verified via RT-PCR experiment. The sequences for B-cell lymphoma 2 gene (Bcl-2), Bcl-2-associated X protein (Bax), PTEN-induced putative kinase 1 (PINK1), beta-actin (β-actin), Peroxisome proliferator-activated receptor gamma coactivator 1 alpha (PGC-1α) and 18S ribosomal RNA (18S) primers are described in Table 1. The RT-PCR conditions were as follows: denaturation (95℃ for 10 min), a three-segment amplification program repeated 30 times (95℃ for 1 min; each annealing temperature for 1 min; 72℃ for 1 min) and final extension (72℃ for 5 min). Electrophoresis was carried out for the PCR products (Mupid-exU, ADVANCE, Japan) in 1.2% agarose gel. Real-time PCR was performed with the cDNA templates. The sequences for caspase 9 (CASP9), caspase 3 (CASP3), Bax, Bcl-2, β-actin and FasR for real-time PCR primers are described in Table 1. Each reaction mixture contained 0.8 µl MgCl2 (final concentration 3 mM), 0.2 µl forward primer (final concentration 0.2 µM), 0.2 µl reverse primer (final concentration 0.2 µM), 1 µl LightCycler DNA Master SYBR Green I 10 X, 1 µl cDNA template and up to 10 µl DW.
RESULTS
Effects on mitochondrial oxidative respiration and copy number
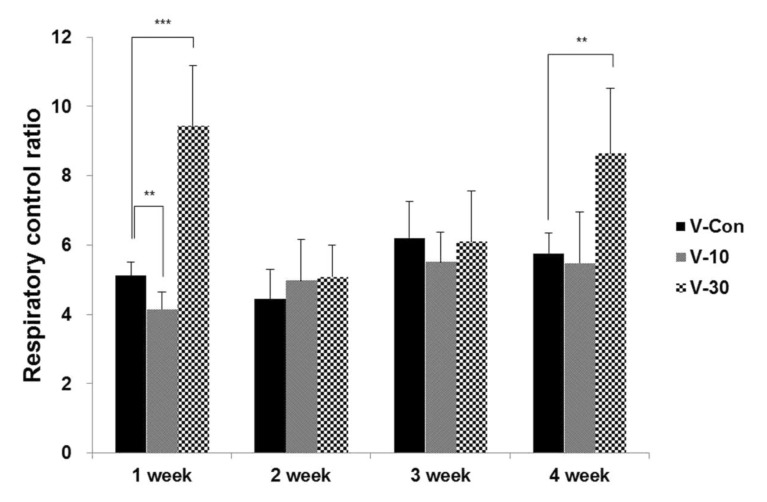
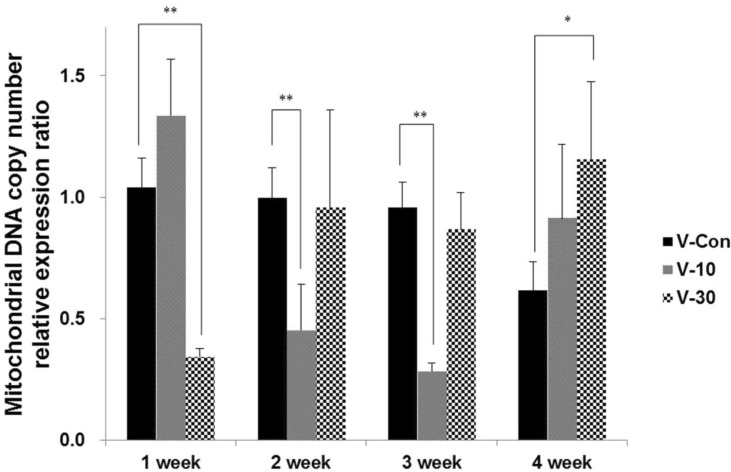
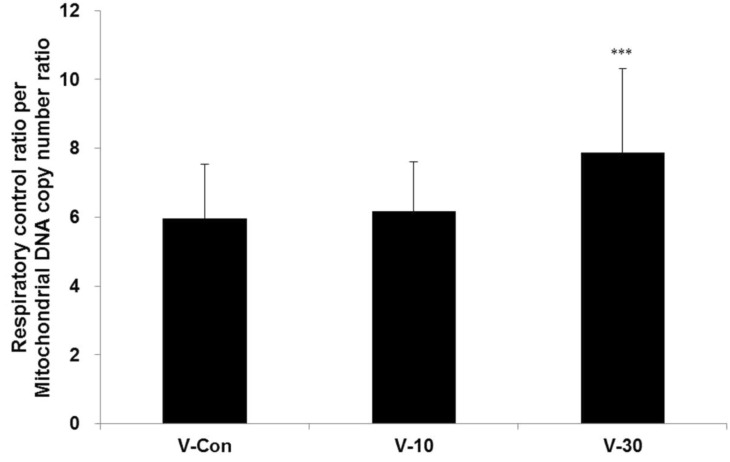
The oxygen consumption significantly increased in the 1st (p<0.001) and 4th (p<0.01) weeks when using the 30% BMCF (Figure 1). The mtDNA copy number significantly decreased with the 30% BMCF in the 1st week (p<0.01) and increased between the 2nd and 4th weeks (Figure 2). The 10% BMCF showed a decrease in the mtDNA copy number between the 2nd and 3rd weeks and a subsequent recovery in the 4th week. However, in the control group, no changes were shown in the mitochondrial oxygen consumption, and the mtDNA copy number slightly decreased as time passed throughout all 4 weeks. The RCR to mtDNA copy number during the 4 weeks showed a dose-dependent increment with the use of the BMCF (p<0.001) (Figure 3). These results suggested that the BMCF invigorate mitophagy and improve mitochondrial oxidative respiration efficiently in skeletal muscle.
Effects on apoptosis- and mitophagy-related genes
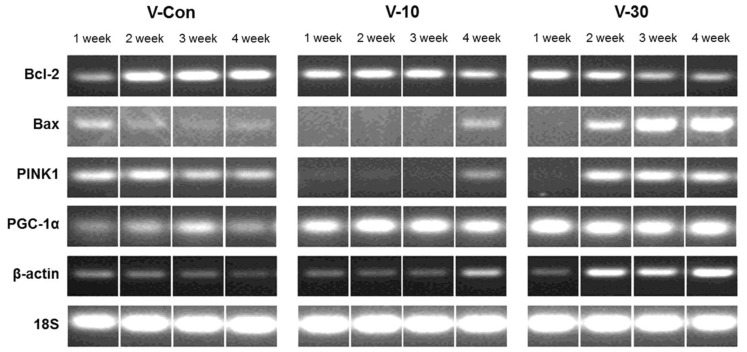
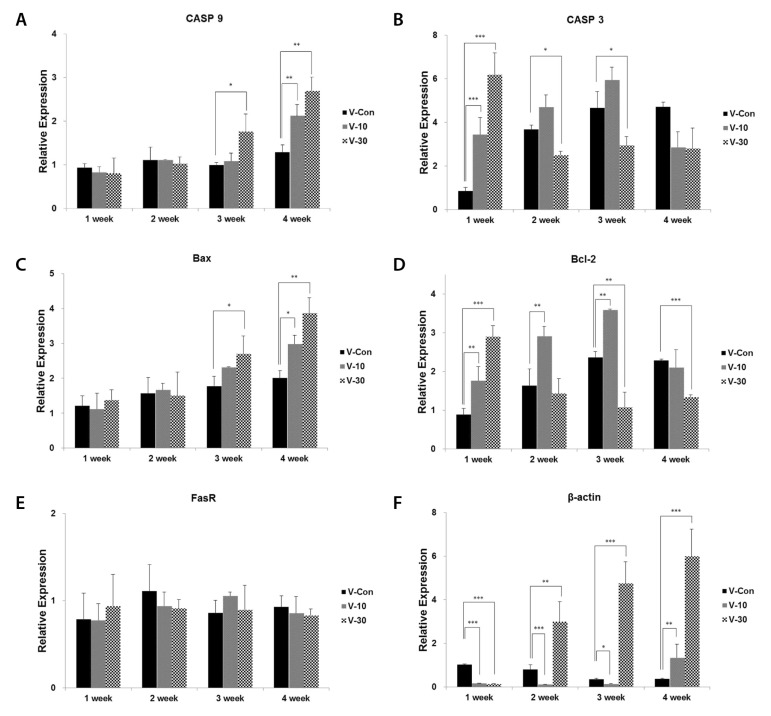
The changes in mRNA expression of Bcl-2, Bax, PINK1 and PGC-1α were confirmed via RT-PCR experiment in a time-course experiment with duration of 4 weeks (Figure 4). In addition, Bcl-2, Bax, CASP3, CASP9 and FasR were confirmed via real-time PCR experiment (Figure 5). In Figure 4, as times passed, Bax expression increased in a dose-dependent manner, although Bcl-2 decreased. PINK1 expression did not change considerably and was inclined to decrease in control group. However, with the use of the BMCF, mRNA expression of PINK1 was down-regulated and subsequently increased in a dose-dependent manner. PGC-1α expression increased alike in 10% and 30% BMCF group compare to control group. β-actin was used as a control marker as 18S, but it also increased with the use of the BMCF. In the results of real-time PCR experiment (Figure 5), the BMCF can be seen to have had no effect on FasR even though Bax and CASP9 expression increased, and also Bcl-2 decreased as former results. CASP3 increased in the 1st week then decreased between the 2nd and 4th weeks in V-30 group, and increased till the 3rd week then decreased in the 4th week in V-10 group, which showing dose-dependent manner. The increment in β-actin due to the use of the BMCF was re-confirmed via real-time PCR with different primer sets. The results of mRNA expression analysis suggest that altered mtDNA copy number is closely related to mitochondrial autophagy and the apoptosis induced by the BMCF is not related to extrinsic death-receptor mediated but mitochondria-mediated signaling pathway.
DISCUSSION
The results of current investigation demonstrated that BMCF provided by VENTEX leads to the stimulation of mitochondrial biogenesis in a dose-dependent manner as evidenced by: increased oxidative respiration of mitochondria, decrement followed by a recovery of mtDNA copy number, improved RCR to mtDNA copy number, increased Bax, CASP9, and PGC-1α, and decreased Bcl-2 expression.
Including mitochondrial mass/volume, mtDNA copy number and mitochondrial respiration are markedly increased at the beginning of myogenic differentiation [26,27,28]. In this study, by wearing the BMCF, the oxidative respiration significantly increased and the mtDNA copy number markedly decreased then recovered as times passed (Figure 2), and RCR to mtDNA copy number during the 4 weeks showed a dose-dependent increment (Figure 3). These results suggest that the BMCF invigorates mitochondrial biogenesis and leads to improvement of mitochondrial oxidative respiration in skeletal muscle.
Bcl-2 and Bax are inversely related to fluctuation of apoptosis and adjust release of cyt c which is known as critical trigger to induce apoptosis signaling cascade [29,30]. Activated Bax induces intrinsic apoptosis signaling pathway using mitochondria intermembrane space generates apoptosis by releasing cyt c to outside, although Bcl-2 blocks the release of cyt c from mitochondria. Also, Bcl-2 plays crucial roles in mitochondria membrane permeability transition with Bax. Decrease in Bcl-2 function and increase in Bax protein levels may be responsible for the release of cyt c from mitochondria [31]. In this study, the mRNA expression of Bax increased in a dose-dependent manner although Bcl-2 decreased, which suggested the BMCF has the facilitating role in mitochondria-mediated intrinsic apoptosis signaling pathway (Figure 4, 5C&D).
PGC-1α co-activates the transcription factors that control expression of genes encoding mitochondrial proteins and, thus, stimulates mitochondrial biogenesis [32,33,34]. In this study, the BMCF upregulated the mRNA expression level of PGC-1α (Figure 4). There are several ways to induce mitophagy in mammalian cells. The PINK1 and Parkin pathway is, so far, the best characterized. This pathway starts in by distinguishing the difference between healthy mitochondria and damaged one. PINK1 has been implicated to detect mitochondria quality [35]. In this study, PINK1 expression did not change considerably and was inclined to decrease in control group, but the expression was down-regulated then subsequently increased with the use of the BMCF in a dose-dependent manner (Figure 4). The result suggests that PINK1-mediated mitochondrial quality control pathway has been triggered by using the BMCF, because the alteration of PINK1 expression and mtDNA copy number showed similar tendency. The mRNA expression level of CASP3 increased in the 1st week then decreased between the 2nd and 4th weeks in V-30 group, and increased till the 3rd week then decreased in the 4th week in V-10 group, which showing dose-dependent manner, which showed diametrically opposite result from PINK1 (Figure 5B). Taken together, the BMCF activated PINK1-mediated mitochondrial quality control pathway and this damaged mitochondrial-targeted PINK1 inhibited apoptosis and triggered mitophagy such as previously reported [36].
Activation of CASP3 is induced by the initiator CASPs, CASP8 and CASP9, as part of the extrinsic and intrinsic apoptotic signaling pathways. In the extrinsic pathway, death ligands, including Fas ligand, bind and activate their cognate receptors which eventually activate CASP8 [37]. In the results of real-time PCR experiment, the BMCF can be seen to have had no effect on FasR even though Bax and CASP9 increased, and also Bcl-2 decreased (Figure 5). These results suggested that the apoptotic effects of BMCF is not relevant to the extrinsic but intrinsic apoptotic signaling pathway, although the verification of CASP8 activity should be left in a subject for further study.
Actin is one of the major components of the cellular scaffold that is necessary for organizing and maintaining cell shape. β-actin dynamics support a many of processes ranging from cell motility, cell division and cell morphogenesis to intracellular progress [38]. In this study, mtDNA copy number has been related to β-actin with similar expression patterns (Figure 4, Figure 5F), which suggested that alteration in the process of mitochondrial biogenesis and oxidative respiration, which is induced by the BMCF, demand sufficient β-actin.
Mitophagy is the selective engulfment of mitochondria by autophagosomes and their subsequent catabolism by lysosomes [35]. It was initially found that mitochondria are selectively engulfed by autophagosomes following a loss in membrane potential [39], suggesting that mitophagy mediates selective removal of damaged mitochondria. In this study, not a few experimental evidences suggested the mitophagy phenomenon. However, the verification of mitochondrial morphology and membrane potential affected by the BMCF is remained.
In summary, the results suggest that the BMCF provided by VENTEX activates PINK1-mediated mitochondrial quality control pathway and mitochondrial biogenesis, which consequently increases the efficiency of oxidative respiration in skeletal muscle. On the other hand, in early stage of apoptosis induced by the BMCF is not related to extrinsic death-receptor mediated but mitochondria-mediated signaling pathway.

 XML Download
XML Download