

 PDF
PDF ePub
ePub Citation
Citation Print
Print


ABBREVIATIONS
DOPA
3,4-dihydroxyphenylalanine
ERK
extracellular signal-regulated kinase
GSK3β
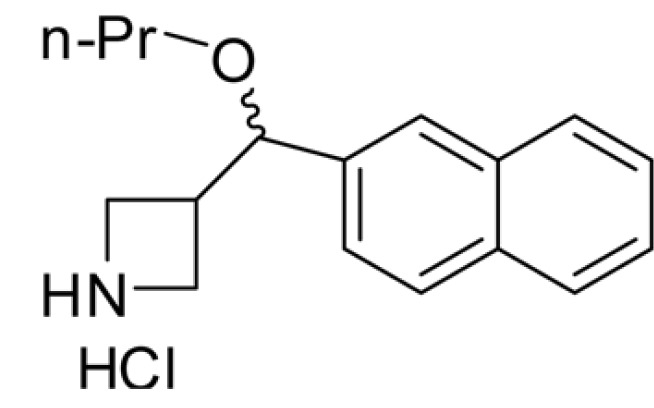
glycogen synthase kinase 3β KHG26792, 3-(naphthalen-2-yl(propoxy)methyl)azetidine hydrochloride
MC1R
melanocortin 1 receptor
MITF
microphthalmia-associated transcription factor
α-MSH
α-melanocyte stimulating hormone
PVDF
polyvinylidene fluoride
TYRP
tyrosinase-related protein
UV
ultraviolet
INTRODUCTION
Skin, the single largest organ, protects the body from DNA-damaging environmental UV radiation by synthesizing melanin [1,2]. Specifically, melanin is synthesized in melanosomes before being transferred to keratinocytes. This process results in skin pigmentation, which is determined by the amount and distribution of melanin [3]. Although melanin protects the skin from UV radiation, over-expression and accumulation of melanin can cause hyperpigmentary skin diseases [4]. As skin whitening is a popular cosmetic issue in Asia [5], novel skin whitening agents are always in development. In a primary screening, we found that KHG26792, 3-(naphthalen-2-yl(propoxy) methyl) azetidine hydrochloride, is an effective skin whitening agent.
Three major enzymes are involved in melanogenesis: tyrosinase, tyrosinase-related protein 1 (TYRP-1), and TYRP-2 [6]. Microphthalmia-associated transcription factor (MITF) stimulates tyrosinase expression [7]. Tyrosinase then mediates the hydroxylation of monophenol and the oxidation of 3,4-dihydroxyphenylalanine (DOPA) into dopaquinone, the major rate-limiting reaction in melanogenesis [8]. Accordingly, inhibition of tyrosinase is a key factor in the regulation of melanogenesis.
In addition to direct inhibition of tyrosinase, tyrosinase expression can be regulated via a number of signal transduction pathways [3,8]. Specifically, extracellular signal-regulated kinase (ERK) represents one major signaling pathway involved in melanogenesis [9,10]. If ERK is phosphorylated and activated, MITF and tyrosinase are down-regulated and melanogenesis is inhibited [9,10].
Because the sale of cosmetics containing agents tested in experimental rodents is prohibited in the European Union, in vitro models are now required to replace animal studies for all products sold in that region [11]. Recently, we reported our skin equivalent (SE) model containing Cervi cornus Colla (deer antler glue) as an alternative for animal skin toxicity testing [12].
In the present study, we established a melanocyte-containing SE model to examine the effects of potential hypopigmentary substances. We investigated the effects of KHG26792 on melanogenesis and related signal transduction pathways, and verified the effects of KHG26792 using our SE model.
METHODS
Drugs
KHG26792, 3-(naphthalen-2-yl(propoxy)methyl)azetidine hydrochloride, was synthesized at the Organic Chemistry Laboratory (KIST, Seoul, Korea) [13], dissolved in DMSO, and stored at -20℃ as a stock solution (50 mM). Dulbecco's Modified Eagle's Medium (DMEM), Nutrient Mixture F-12 Ham, sodium bicarbonate, HEPES, formaldehyde, insulin, L-ascorbic acid, isoproterenol, hydrocortisone, 12-O-tetradecanoylphorbol-13-acetate (TPA), cholera toxin (CT), synthetic melanin, L-DOPA, and mushroom tyrosinase were obtained from Sigma (St Louis, MO, USA). DMEM/Nutrient Mixture F-12, (3:1, DMEM:F-12), antibiotic-antimycotic (penicillin, streptomycin), trypsin-EDTA, and sodium hydroxide were purchased from WelGENE (Dalseogu, Daegu, South Korea). Cervi cornu Colla (CCC) was obtained from Siwon Herbal Medicine Co. (Jincheon-gun, South Korea), hyaluronic acid (HA) was from LG Life Sciences, Ltd. (South Korea), fetal bovine serum (FBS) was from Hyclone (Logan, UT, USA), and recombinant human epidermal growth factor (EGF) was from Invitrogen Co. (Gibco, Camarillo, CA). Antibodies specific for phospho-ERK1/2 (Thr202/Tyr204, #9101S), total (phosphorylated and nonphosphorylated) ERK1/2 (#9102), phospho-GSK3β (#9336), and phosphorylated Akt (Ser473, #9271) were purchased from Cell Signaling Technology (Beverly, MA, USA). Antibodies specific for tyrosinase (C-19) and actin (I-19) were purchased from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA, USA). Microphthalmia Ab-1 (C5, MS-771-P0) was obtained from NeoMarkers (Fremont, CA, USA). Secondary antibodies specific for anti-goat IgG (PI-9500), antimouse IgG (PI-2000), and anti-rabbit IgG (PI-1000) were purchased from Vector Laboratories (Burlingame, CA, USA).
Cell culture
The Mel-Ab cell line is a spontaneously immortalized murine melanocyte cell line that produces large amounts of melanin [14]. Mel-Ab cells were incubated in DMEM supplemented with 10% (v/v) FBS, 100 nM TPA, 1 nM CT, 50 µg/ml streptomycin, and 50 µg/ml penicillin at 37℃ in 5% CO2. CCD-25Sk human fibroblasts were purchased from American Type Culture Collection (ATCC, Rockville, MD, USA). HaCaT human keratinocytes [15] were purchased from Cell Lines Service (Eppelheim, Germany). Cells were grown in DMEM supplemented with 10% (v/v) FBS, 50 µg/ml streptomycin, and 50 µg/ml penicillin at 37℃ in 5% CO2.
Cell viability assay
Cell viability was determined using a crystal violet assay. Mel-Ab cells were treated with KHG26792 for 24 h, the medium was removed, and the cells were stained with 0.1% crystal violet in 10% ethanol for 5 min at room temperature. The cells were rinsed four times with distilled water and the crystal violet retained by adherent cells was extracted with 95% ethanol. Absorbance was determined at 590 nm using an ELISA reader (VERSAMax; Molecular Devices, Sunnyvale, CA, USA).
Measurement of melanin content and microscopy
Melanin content was measured using a previously described method, with slight modifications [16]. Cells were treated with various concentrations (0.1~20 µM) of KHG26792 in DMEM containing 10% FBS for 3 days. Cell pellets were then dissolved in 550 µl of 1 N NaOH at 100℃ for 30 min and centrifuged at 13,000×g for 5 min. The resulting supernatants were analyzed at 400 nm using an ELISA reader. Before measurement of melanin content the cells were observed using a phase contrast microscope (Olympus IX50, Tokyo, Japan), and photographed using a DCM300 digital camera (Scopetek, Inc., Hangzhou, China) supported by ScopePhoto software (Scopetek, Inc.).
Tyrosinase activity
A cell-free assay system was used to determine the direct effect of KHG26792 on tyrosinase activity. Briefly, 70 µl of phosphate buffer containing KHG26792 was mixed with 20 µl of mushroom tyrosinase (stock solution 53.7 units/ml) and 10 µl of 10 mM L-DOPA. After incubation at 37℃ for 20 min, the absorbance was measured at 475 nm.
Western blot analysis
Mel-Ab cells were lysed in cell lysis buffer (62.5 mM Tris-HCI [pH 6.8], 2% SDS, 5% β-mercaptoethanol, 2 mM phenylmethylsulfonyl fluoride, protease inhibitors [Complete™; Roche, Mannheim, Germany], 1 mM Na3VO4, 50 mM NaF, and 10 mM EDTA). Proteins in the total cell lysates were separated by SDS-polyacrylamide gel electrophoresis using 20 µg of protein per lane, blotted onto polyvinylidene fluoride (PVDF) membranes, and blocked with 5% dried milk in Tris-buffered saline containing 0.5% Tween 20. Blots were incubated with the appropriate primary antibodies at a dilution of 1:1000 and then with horseradish peroxidase-conjugated secondary antibody. Bound antibody was identified using an enhanced chemiluminescence testing kit (Thermo Scientific Inc., Bremen, Germany). All immunoblot images were obtained using a LAS-1000 lumino-image analyzer (Fuji Film, Tokyo, Japan).
Preparation of dermal equivalents
Dermal equivalents were prepared as previously described with some modifications [17]. In brief, type 1 collagen extracted from rat tail tendons was dissolved by stirring in 1/1000 glacial acetic acid at 4℃ for 48 h. Dermal equivalents were made by mixing the following solutions: 1% type 1 collagen solution, 10×medium (DMEM:Nutrient Mixture F-12 Han=3:1), 10×reconstitution buffer (0.05 N NaOH, 0.26 mM NaHCO3, 200 mM HEPES), 5 mg/ml hyaluronic acid (HA), and 0.33 g/ml CCC. Dermal equivalents including CCC were prepared by composition, as described in a previous study [12]. All dermal equivalents contained 3×105 fibroblasts. Three milliliters of this mixture was transferred into 24-mm transwell inserts with a 3.0 µm pore polycarbonate membrane (Corning, Inc., USA), which was then placed in the incubator at 37℃ for gelation.
Reconstruction of pigmented skin equivalents
To reconstruct pigmented SEs, dermal equivalents were treated with 0.03 mg/ml collagen type IV and 1×105 Mel-Ab cells were seeded onto the dermal equivalents. HaCaT cells (1×106 cells) were seeded onto the dermal equivalents and cultured submerged for 1 day, and then at the air-liquid interface for 12 days. The growth medium was DMEM/F-12 supplemented with 5% FBS, 0.4 µg/ml hydrocortisone, 1 M isoproterenol, 25 µg/ml ascorbic acid, and 5 µg/ml insulin. For the submerged culture, 1 ng/ml epidermal growth factor (EGF) was added to the growth medium, whereas 10 ng/ml EGF also was added for the air-liquid interface culture. The medium was changed three times per week. Every time the medium was changed, 10 or 50 µM KHG26792 was added to the new solutions.
Immunohistochemistry and Fontana-Masson stain assay
After 13 days, the SEs were fixed in 10% formaldehyde for 1 day, embedded in paraffin, and 4- to 6-µm sections were prepared. For morphologic analysis, sections were stained with hematoxylin and eosin (H&E). To detect melanin, a Fontana-Masson stain kit (ScyTek Laboratories, Logan, UT, USA) was used according to the manufacturer's instructions.
RESULTS
KHG26792 decreased melanin synthesis in Mel-Ab cells
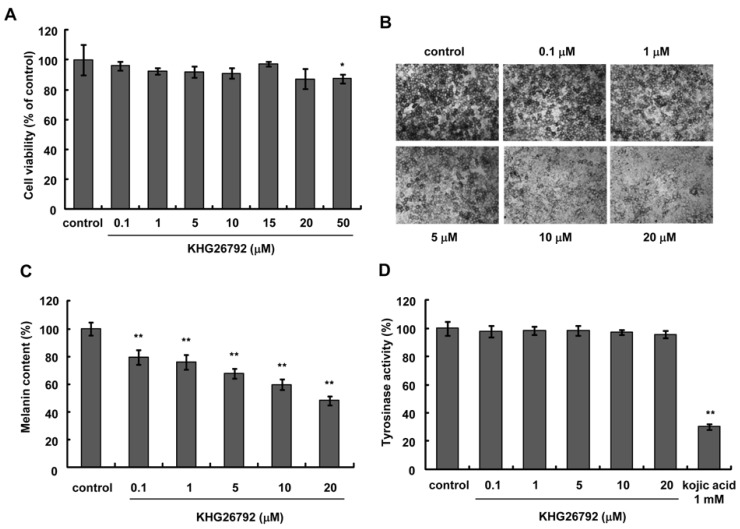
The structure of KHG26792 is displayed in Fig. 1. To evaluate the safety of KHG26792 in Mel-Ab cells, cells were treated with different concentrations of KHG26792 (0.1~50 µM) and cell viability was measured using a crystal violet assay. As shown in Fig. 2A, KHG26792 had no cytotoxic effects at concentrations between 0.1 and 20 µM. Higher concentrations of KHG26792 (50 µM) were found to be slightly cytotoxic compared with the untreated control. Therefore, concentrations between 0.1 and 20 µM were used in the following experiments. To examine the effects of KHG26792 on melanogenesis, cells were incubated with KHG26792 (0.1~20 µM) for 3 days. As shown in Fig. 2B, cell morphology indicated that the amount of melanin decreased after treatment with KHG26792 in a dose-dependent manner. Measurement of melanin content confirmed that KHG26792 decreased the amount of melanin in a dose-dependent manner (Fig. 2C). As tyrosinase is the key enzyme in melanin synthesis [18], tyrosinase activity was measured in a cell-free system after the addition of KHG26792. The results indicated that KHG26792 did not block tyrosinase directly (Fig. 2D).
KHG26792 suppressed MITF and tyrosinase expression via the ERK pathway
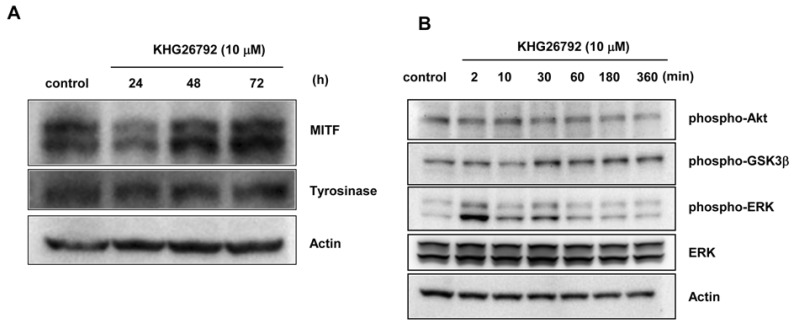
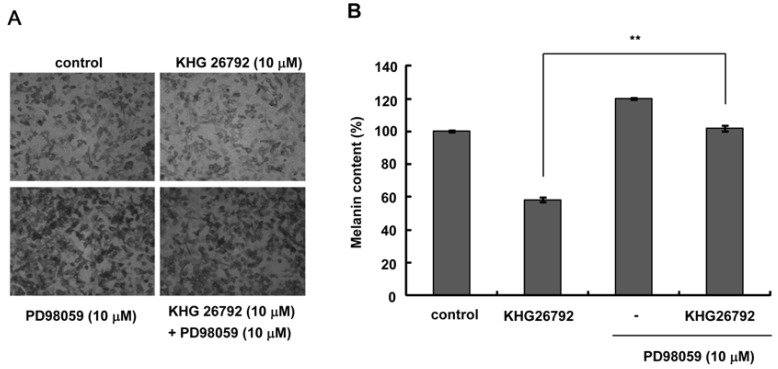
Because KHG26792 did not inhibit tyrosinase directly, the influence of KHG26792 on the expression of melanogenic proteins was analyzed by Western blot analysis, which showed that KHG26792 downregulated MITF and tyrosinase at 24 h (Fig. 3A). Decreased expression of MITF and tyrosinase was also observed at 48 h. We next investigated the status of melanogenesis-associated signal transduction pathways after treatment with 10 µM KHG26792. The resulting data indicate induction of ERK phosphorylation, which peaked at 2 min and lasted for a minimum of 30 min, whereas phosphorylation of Akt and GSK3β did not change (Fig. 3B). It is well known that activating the ERK pathway results in downregulation of MITF and decreased melanin synthesis, therefore we measured melanin content in the presence of PD98059, an ERK pathway inhibitor. As shown in the photographs in Fig. 4A, PD98059 rescued the KHG26792-induced hypopigmentation and decreased melanin content resulting from treatment with KHG26792 (Fig. 4B).
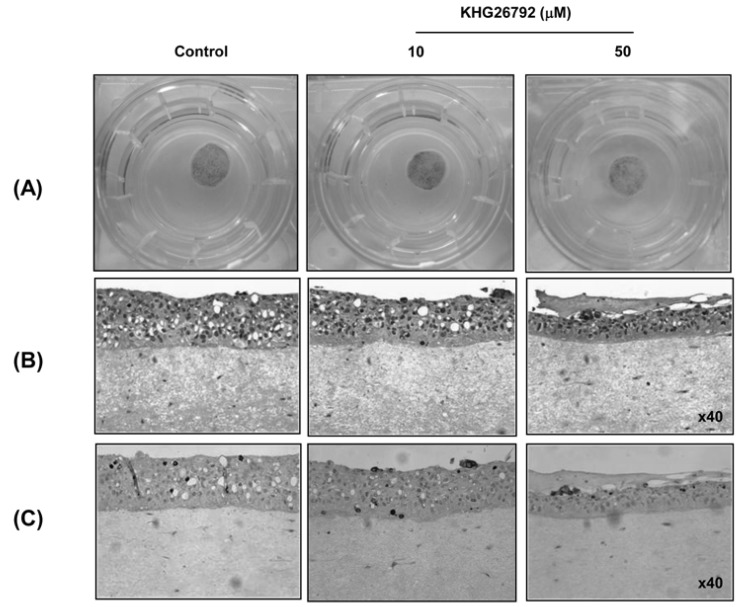
KHG26792 inhibited melanogenesis in a pigmented skin equivalent model
The pigmented SE was prepared as described in Methods. The color of the skin equivalent treated with 50µM KHG26792 was lighter than that of the untreated control, whereas treatment with 10 µM KHG26792 yielded no change (Fig. 5A). As shown in Fig. 5C, melanin staining was reduced by treatment with 50 µM KHG26792.
DISCUSSION
Abnormal melanin accumulation may result in hyperpigmentation and other cosmetic problems in humans [6]. Therefore, new therapeutic agents that prevent melanin synthesis are currently being developed for skin whitening [19,20]. In the present study, we found that KHG26792 significantly reduced melanin synthesis in Mel-Ab cells and a pigmented skin equivalent.
MITF and tyrosinase are key factors in melanogenesis. Many kinds of whitening agents, notably kojic acid and arbutin, reduce melanin synthesis by directly inhibiting tyrosinase activity [21]. However, KHG26792 did not directly affect the activity of tyrosinase (Fig. 2D), but instead reduced expression of MITF and tyrosinase at 24 h after treatment. ERK activation has previously been reported to reduce melanin synthesis by decreasing MITF expression [8,22]. We found that KHG26792 phosphorylated and activated ERK. Furthermore, a specific inhibitor of the ERK pathway, PD98059, rescued the decreased melanin synthesis resulting from treatment with KHG26792 (Fig. 4). Based on these data, we propose that KHG26792-mediated ERK activation results in the inhibition of melanin synthesis.
Two primary testing methods exist to assess the effects of medications or cosmetics: in vitro and in vivo methods. Recently, the sale of cosmetics tested using animal experimentation was prohibited in the European Union. An SE represents a good alternative to animal studies [11]. In vitro models (including SEs) that possess a similar structure to normal human skin have been developed over the past 30 years [23,24]. Many studies have demonstrated that paracrine factors secreted from keratinocytes and fibroblasts can affect melanogenesis in melanocytes [25,26]. Therefore, skin whitening agents can clearly be evaluated using a pigmented SE model [27]. We recently improved our SE model using Cervi cornus Colla (deer antler glue) [12]. In the present study, we constructed pigmented SEs that successfully simulated the color of skin (Fig. 5).
An important aspect of the present study is that KHG26792 was proven via Fontana-Masson staining to have whitening effects in our pigmented SE model (Fig. 5C). To establish an SE as an easy and economical test model, we used three different cell lines: epidermal HaCaT cells, dermal CCD-25Sk cells, and Mel-Ab melanocytes. The location of melanin in the SE is different from that in real human skin, where melanocytes are located in the basal layer of the epidermis [28]. However, it was technically difficult to maintain melanocytes at the basal layer, and pigmented skin equivalents could be improved by further studies.
In conclusion, KHG26792 exhibited skin whitening effects both in a monolayer cell culture model and a pigmented skin equivalent model, most likely through the activation of ERK. Moreover, the pigmented skin equivalents developed in this study could be used as a general model for testing skin whitening agents.
 XML Download
XML Download