

 PDF
PDF ePub
ePub Citation
Citation Print
Print


ABBREVIATIONS
CAT
catalase
DCF-DA
2',7'-dichloro-fluorescein diacetate
COX-2
cyclooxygenase-2
DMEM
Dulbecco's modified Eagle's medium
EEC
esophageal epithelial cells
HBSS
Hank's Balanced Salt Solution-Modified
FBS
fetal bovine serum
GAPDH
glyceraldehyde-3-phosphate dehydrogenase
IκB
inhibitor-κB
MTT
dimethyl thiazolyl diphenyl tetrazolium salt
NF-κB
nuclear factor kappa-light-chain-enhancer of activated B cells
PBS
phosphate-buffered saline
PGE2
prostaglandin E2
QGC
quercetin-3-O-β-D-glucuronopyranoside
ROS
reactive oxygen species
SOD
superoxide dismutase
INTRODUCTION
Reactive oxygen species (ROS) participate and regulate diverse downstream signaling pathways leading to specific cellular functions [1,2] such as growth, metabolic rate, cell division, necrosis, apoptosis and aging processes [3-5]. But the imbalance between formation and neutralization of ROS causes oxidative stress [6]. The oxidative stress causes toxic effects through the production of nitric oxide, peroxynitrite, superoxide anion, hydrogen peroxide, hypochlorous acid as well as hydroxyl radical that damage all components of the cells including proteins, lipids, and DNA [5,7]. Large amounts of ROS have been reported to be involved in the etiology of several human diseases, including inflammation, cardiovascular and neurodegenerative disorders and cancer [1,8]. Increased mucosal concentrations of ROS have also been implicated in the generation of gastrointestinal diseases, including the acid peptic diseases and the inflammatory disorders [9].
Inflammation is one of the complex biological responses to harmful stimuli such as ROS, NO, cytokines, microbial products, and UV [10]. In fact, inflammation, which is functionally a protective response, can be considered by the organism to remove the injurious stimuli as well as initiate the healing process for the tissue. However, chemical mediators of inflammation such as histamine, cytokines and prostaglandins can lead to a number of diseases [11]. Therefore, inhibition of these mediators can decrease the inflammatory response.
NF-κB is a protein transcription factor that functions to enhance the transcription of a variety of genes, including cytokines and growth factors, adhesion molecules, immunoreceptors and acute-phase proteins [12]. ROS can activate NF-κB by stimulating IκB degradation in a variety of cell types. NF-κB activation could play a role in cyclooxygenase-2 (COX2) regulation and increased secretion of prostaglandin E2 (PGE2) which modulates cytokine balance, inhibits host immunity and plays an important role in inflammation [13].
Cell damage caused by free radicals in gastric or esophageal mucosa can be prevented by administration of free radical scavengers [14]. A major endogenous antioxidant in cells is the enzyme superoxide dismutase (SOD), which catalyzes the superoxide anion into hydrogen peroxide and molecular oxygen [15]. SOD is the first enzymatic step that protects cells from toxic ROS [16]. Catalase is the next enzymatic step that converts hydrogen peroxide to water and molecular oxygen [17]. Previous research demonstrated that antioxidant capacity, as determined by the measurement of SOD activity, is depressed in esophagitis and this depression is significant in severe esophagitis [18]. Animal studies have shown that reflux esophagitis in rats is mediated by oxygen-derived free radicals, and that SOD provides an important protective effect against reflux-induced esophageal injury [19].
Flavonoids are considered relatively non-toxic bioactive substances and display diverse biological effects such as anti-inflammatory, anti-oxidative, anti-allergic, hepato-protective, anti-thrombotic, anti-viral, and anti-carcinogenic activities [20]. Of the many actions of flavonoids, antioxidant and anti-inflammatory effects stand out. The best-described property of almost every group of flavonoids is their capacity to act as antioxidants, and the inhibitory activity on inflammatory cells appears to suppress any other clinically available compound [21].
Many genes and signal pathways are influenced by ROS and antioxidants [22]. Quercetin is a potent natural antioxidant that attenuates H2O2-induced cytotoxicity [23] and prevents the gastric mucosal lesions produced by ethanol [24]. Quercetin-3-O-β-D-glucuronopyranoside (QGC) is a flavonoid glycoside extracted from Rumex Aquaticus. In our preliminary study, QGC is more potent than quercetin on inhibition of experimental reflux esophagitis and indomethacin-induced gastritis in S.D rats, in vivo (not published). The efficacies of QGC were caused by decreasing ulcer index, gastric volume, total acidity, TBARS.
In this study, the modulating effects of QGC in feline esophageal epithelial cells (EECs) on cell damage induced by a variety of inflammatory inducers, such as acid, LPS, indomethacin and ethanol, were investigated in vitro. Especially, we focused on the anti-oxidative and anti-inflammatory effects of QGC on acid-induced cell injury since acid plays a significant role in the development of gastroesophageal reflux symptoms [25]. Data show that QGC obviously has cytoprotective effects by antioxidant and anti-inflammatory ability in EECs.
METHODS
Materials
QGC was supplied by Pharmacal Botany Resources Laboratory (Chung-Ang Univ., Seoul, Korea). NF-κB antibody from Cell Signaling (Beverly, MA); SOD-2, Catalase, GAPDH and lamin B antibody from Santa Cruz Biotechnology (Santa Cruz, CA); COX2 antibody from BD Biosciences (Billerica, MA); thiazolyl blue tetrazolium bromide, LPS, indomethacin, 2',7'-Dichloro-fluorescein diacetate (DCF-DA) and SOD assay kit from Sigma Chemical Co. (St Louis, MO), catalase assay kit and PGE2 Express EIA kit from Cayman chemical (Ann arbor, MI).
Preparation of cat esophageal epithelial tissue squares and cell culture
All animal experiments were performed in accordance with the 'Principles of laboratory animal care' of NIH guidelines. Adult cats of either sex weighing between 2.5 and 4.0 kg were anesthetized with zoletil 50 (12.5 mg/0.25 ml/kg). Esophageal epithelial tissue squares were then prepared as described previously [26].
The sliced tissue was then placed into DMEM supplemented with 10% FBS containing antibiotics and incubated in a humidified atmosphere of 5% CO2 and 95% air at 37℃. After 10 days later, the medium was exchanged with fresh 10% FBS-DMEM. After reaching confluence, cells were detached with 1% trypsin in HBSS with bicarbonate. Cells were then seeded at 2×105 cells/ml on 100-mm culture dishes, and maintained in 10% FBS-DMEM. The medium was changed every 48 h until the cells reached confluence. We confirmed typical epithelial morphology of the primary-cultured cells and identified those cells by an indirect immunofluorescent staining method using a cytokeratin monoclonal antibody [26]. Experiments were performed on cells of passage 3 or 4.
Measurement of cell viability
Cell viability was determined by the conventional MTT reduction assay. Viable cells convert MTT to insoluble blue formazan crystals by mitochondrial respiratory chain enzyme succinate dehydrogenase. Cells were seeded at a density of 2×104/6-well plates and maintained in 10% FBS-DMEM. They were made quiescent at confluence by incubation in serum-free DMEM for 24 h, followed by treatment with the given concentrations of each compound for the indicated time. Cells were then washed with PBS and treated with MTT solution (final concentration, 5 mg/ml) for 4 h at 37℃. The supernatant was removed and the formazan crystals were dissolved with 500 µl DMSO. Absorbance at 570 nm was measured with a microplate reader (Molecular Devices, Sunnylvale, CA).
Measurement of ROS production
DCF-DA was used to measure levels of intracellular H2O2 production induced by acid, LPS, indomethacin and ethanol [27]. Cells were grown on 6-well plates for 2 day and starved in serum-free DMEM for 24 h. The cells were treated with 50 µM QGC 1 h and then incubated with H2O2-sensitive fluorophore DCF-DA (20 µM) for 1 h at 37℃ in dark. The cells were washed with Krebs solution and exposed to acid, LPS, indomethacin or ethanol for the indicated time at 37℃ in dark. The cells were harvested and centrifuged for 5 min at 10,000 rpm at 4℃. After the pellets were resuspended with Krebs buffer, DCF fluorescence was measured by a fluorospectrophotometer using excitation and emission wavelengths of 485 and 535 nm, respectively (Tecan, GENios Pro).
Measurement of SOD-2 and catalase activity
Serum-starved cells were pretreated with 50 µM QGC for the indicated time before incubation with acid for 10 min. After the incubation, cells were harvested with lysis buffer. The lysates were scraped, collected into eppendorf tubes, sonicated (6 sec, 3×) and then centrifuged for 10 min at 13,000 rpm at 4℃, and the supernatants were used for SOD and catalase assays using SOD assay kit and catalase assay kit.
Measurements of PGE2 released from EECs
For experiments designed to measure the release of PGE2 due to endogenous arachidonic acid, the medium was collected, centrifuged, and stored at -70℃ until assay. The level of PGE2 released into cultured medium was quantified using a PGE2 Express EIA kit.
Preparation of cell extracts
When cells reached confluence, they were starved by incubation in serum-free DMEM for 24 h. The cells were stimulated with 50 µM QGC for the indicated time before incubation with acid (pH 4) for 10 min. hey were washed with ice-cold PBS and lysed with ice-cold lysis buffer for 5 min. The lysates were scraped, collected into eppendorf tube, sonicated (6 sec, 3×) and then centrifuged at 13,000 rpm for 10 min at 4℃, and the supernatants were collected.
Preparation of nuclear proteins
The starved cells were incubated with 50 µM QGC for the indicated time before incubation with acid for 10 min. They were washed with PBS and centrifuged at 3,000 rpm for 5 min at 4℃. The pellets were resuspended in ice-cold isotonic buffer A (10 mM HEPES), 10 mM KCl, 0.1 mM EDTA, 1 mM DTT, 0.5 mM PMSF) and protease inhibitor cocktail. After 15 min incubation in an ice bath, cells were vortexed vigorously for 10 sec with addition of 10% NP-40 and recentrifuged at 13,000 rpm for 2 min at 4℃. The pellets were resuspended in ice-cold buffer B containing 20 mM HEPES, 0.4 M NaCl, 1 mM EDTA, 10% glycerol, 1 mM DTT, 1 mM PMSF and protease inhibitor cocktail followed by incubation at 4℃ for 30 min with periodic vortexing. The mixture is then centrifuged at 13,000 rpm for 10 min at 4℃. The supernatant was collected and stored at -70℃ for protein assay and western blotting.
Western blot analysis
Western blot analysis was performed as described previously [26] SOD-2 and catalase expressions, or NF-κB translocation was calculated as the ratios of SOD-2, catalase or COX2 to GAPDH, or NF-κB to lamin B, respectively.
RESULTS
Effect of QGC on cell viability of cultured feline EECs
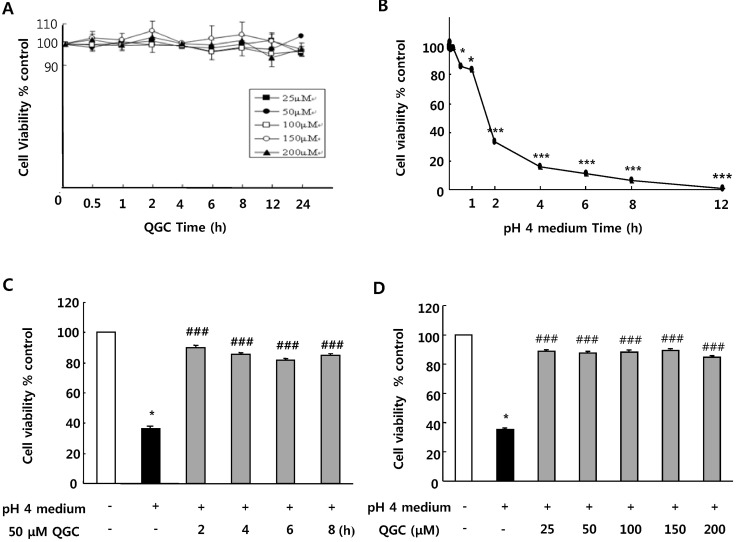
First, we assessed whether QGC itself has any effect on cell viability in cultured EECs. Serum-starved cells were incubated with the given concentration of QGC for the indicated time. When cell viability was measured using MTT assay, there was no detrimental effect of QGC itself (Fig. 1A).
Since acid is critical to the development of esophagitis, the effect of acid exposure time on cell viability was examined by MTT assay (Fig. 1B). There was no significant decrease in cell viability of EECs incubated in pH 4 medium (acid) for 10 min. After 30 min of exposure to acid, cell viability was however decreased to about 86%. In addition, exposure of cells to acid for 2 h led to a significant decrease below 50% in cell viability. Incubation of cells with acid for 12 h caused almost complete cell death.
To investigate protective effect of QGC against acid-induced EECs damage, we performed time-course and concentration-course analyses of its effect using MTT assay. When serum-starved cells were pre-incubated with 50 µM QGC for 2~8 h (Fig. 1C), or with 25~200 µM QGC for 2 h (Fig. 1D), the viability of cells exposed to acid for 2 h significantly recovered to above 80% compared to control, while the acid treatment alone for 2 h caused a significant decrease to about 36% in cell viability. These results suggest that QGC enhanced viability against acid-induced cytotoxicity.
Modulating effect of QGC on intracellular H2O2 level in cultured EECs stimulated by various inducers of inflammation
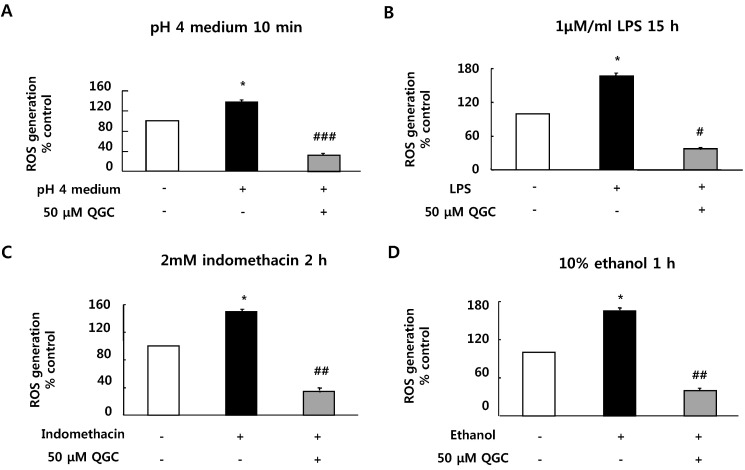
Serum-starved EECs were pre-incubated with the presence of 50 µM QGC for 2 h. Cells were then stimulated with the indicated inflammatory inducers and their ROS production was estimated using DCF-DA assay (Fig. 2). Inflammatory inducers including acid (pH 4 medium, 10 min), LPS (1 µM/ml, 15 h), indomethacin (2 mM, 2 h) and ethanol (10%, 1 h) induced significant increases in ROS production by 138.11%, 166.69%, 149.35% and 164.92% vs. control, respectively. However, when cells were pretreated with 50 µM QGC for 2 h, ROS levels were significantly decreased below about 40% in each group. These results suggest that QGC has ROS scavenging effects in EECs.
Effect of QGC on the activities and expressions of SOD and catalase in EECs exposed to acid
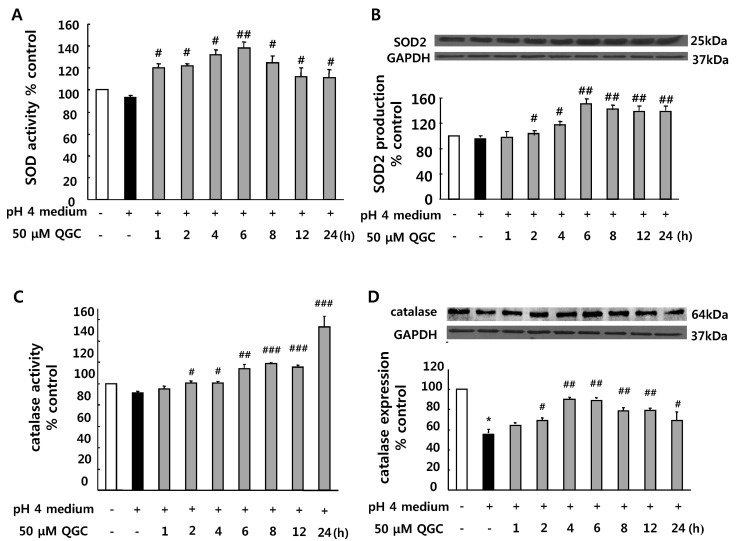
We then examined whether QGC activates ROS scavenging enzymes such as SOD and catalase by EIA kit for assay. Pretreatment of cells with 50 µM QGC for the indicated time before 10 min of exposure to acid significantly increased activities of SOD and catalase. QGC caused SOD activation at 1 h and reached a maximal response at 6 h (Fig. 3A). Catalase was activated at 2 h and the activation was increased in time-dependent manner (Fig. 3C).
Expressions of SOD and catalase were also measured by western blotting (Fig. 3B and 3D). Serum-starved cells were preincubated with 50 µM QGC for the indicated time and then stimulated with acid for 10 min. QGC caused significant increases in SOD expression as compared to the group treated with acid alone. Interestingly, QGC restored the decrease of catalase expression in EECs exposed to acid. The inductions of SOD and catalase expression were detected at 2 h and reached a maximal response at 6 h.
Inhibitory effects of QGC on acid-induced NF-κB translocation in EECs
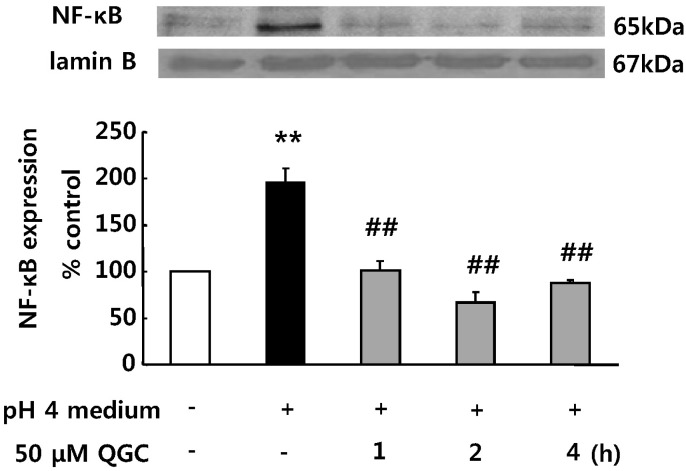
We next investigated whether EECs exposed to acid induces NF-κB translocation which is involved in the induction of inflammatory gene expression, and whether QGC inhibits the NF-κB translocation using western blotting (Fig. 4). When serum-starved cells were stimulated with acid alone, their nuclear extracts exhibited NF-κB translocation in 10 min (195.4% vs. control). Expression level of lamin B was unaffected by acid. However, pretreatment with QGC for the indicated time before exposure to acid for 10 min significantly prevented the NF-κB translocation (67.1% vs. control).
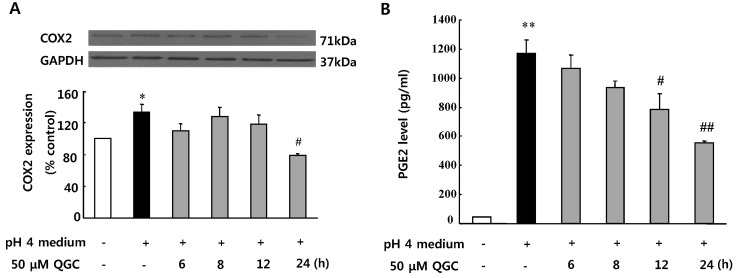
Inhibitory effects of QGC on COX2 expression and PGE2 secretion in EECs exposed to acid in EECs
Effect of QGC on acid-induced COX2 expression was also tested by western blotting (Fig. 5A). Incubation of cells with acid stimulated COX2 expression by 133.8% at 10 min, compared to the control. The acid-induced COX2 expression was significantly inhibited by pretreatment of cells with 50 µM QGC for 24 h.
Therefore, inhibitory effect of QGC on the acid-induced PGE2 synthesis was also determined using PGE2 EIA kit (Fig. 5B). Data show that treatment of EECs with acid caused a significant increase in the PGE2 production at 10 min (1171.92 pg/ml). The acid-induced PGE2 release was decreased by pretreatment of cells with 50 µM QGC in a time-dependent manner. Noticeably when QGC was pretreated for 24 h, the acid-induced PGE2 release was reduced to 555.88 pg/ml. The basal level of PGE2 was 43.98 pg/ml.
DISCUSSION
Esophageal reflux occurs when gastric contents move into the esophagus. Reflux occurs when the lower esophageal sphincter is weak or relaxes. When the gastric contents exposed to esophageal mucosa long time, it would develop into gastroesophageal reflux disease (GERD) [28]. The Agency for healthcare Research and Quality defines GERD as heartburn and/or acid regurgitation occurring weekly [29]. GERD is one of the most important and frequent gastrointestinal disorders, and may lead to esophageal cancer [30]. Reflux of gastric acid into the esophagus is an important factor in esophagitis. In the present study the acidic medium was used to represent gastroesophageal reflux [31].
Experimental studies suggest that free radicals are involved in acid-induced damage of esophageal mucosa [32]. The present study also shows that exposure of cells to acid induced cell injury and significantly produced ROS. In addition, we confirmed that EECs exposed to LPS [33], indomethacin [34], or ethanol [35] known as inflammatory inducers produce intracellular ROS.
Administration of anti-oxidants could attenuate the severity of experimentally induced reflux esophagitis [36] and prevent or treat reflux esophagitis [37]. Flavonoids are reported as free radical scavengers and cytoprotective compound [7,21]. Especially, they exhibited protection against H2O2-mediated cytotoxicity [23]. In the present study, we demonstrated that QGC acts as non-stressful and non-cytotoxic anti-inflammatory flavonoid since QGC exhibits protective effect against acid-induced cell damage. Our data also suggest that QGC acts as a potent ROS scavenger in EECs since overproduction of intracellular ROS generated by a variety of inflammatory inducers were significantly inhibited by QGC.
Overproduction of ROS has been implicated in the generation of esophagitis by depression of MnSOD activity [9,18,19]. MnSOD (SOD2) is considered a protective enzymatic protein for a number of insults including esophageal refluxinduced oxidative damage: Overexpression of the MnSOD in the esophagus can prevent irradiation-induced esophagitis in the mouse model [38]; In rat models, decreases in MnSOD expression and activity are associated with esophagitis and cell death [19]. Interestingly, flavonoids induce SOD expression in various cell types [39], and elevate catalase activity [40]. In this study, QGC up-regulated the decreased SOD and catalase activity and expression in EECs exposed to acid. Thus, our data imply that the ROS-scavenging capacity of QGC is derived from the enhancement of SOD and catalase expressions.
ROS can activate NF-κB by stimulating IkB degradation in a variety of cell types [16]. Quercetin prevents oxidative stress through NF-κB inactivation in gastric mucosa of rats [20], and suppresses pro-inflammatory cytokines production through NF-κB pathway [23]. Our data show that QGC inhibited acid-induced NF-κB nuclear translocation, suggesting anti-inflammatory activity of QGC through the suppression of NF-κB translocation.
Although not fully understood, several cellular action mechanisms are proposed to explain the anti-inflammatory action of flavonoids [41]. Our data demonstrate the modulating effects of QGC on COX2 expression and PGE2 secretion in EECs stimulated with acid. The data also show that PGE2 secretion is in parallel with COX2 expression, indicating that PGE2 secretion is mediated through the enhanced expression of COX2. Our findings are consistent with the previous observation that a natural antioxidant such as chebulagic acid showed a potent anti-inflammatory effects which are exerted via inhibition of PGE2 production and down-regulation of COX2 through the blockade of ROS generation and NF-kB activation in LPS-stimulated phagocytes [42].
In conclusion, our data suggest that QGC exhibits potent down regulation of ROS generation and protective activity against acid-induced cell damage in EECs. QGC exerts the anti-oxidative effect through the expression of SOD and catalase as well as their activation. Possible cellular mechanism of anti-inflammatory action of QGC is an inhibition of NF-κB translocation, COX2 expression and PGE2 secretion. These experiments demonstrate that QGC could be a promising drug for the prevention of reflux esophagitis and a scavenger of ROS generated by inflammatory inducers.
 XML Download
XML Download