

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Vestibular system controls blood pressure and respiration by vestibulo-autonomic reflex through some interneurons [1]. Electrical or selective natural stimulation of the vestibular receptors in cats elicits an increase in sympathetic nerve activity [2,3]. A large amount of evidence suggests that vestibulo-sympathetic reflex pathways are independent of baroreceptor reflex pathways in various species [4-6]. The vestibular system contributes to the pressor response in conscious rats during exposure to gravitational changes [7,8]. Doba and Reis [9] were the first to clearly demonstrate that the vestibular system plays a role in cardiovascular responses to postural changes. A postural change from a lying position to a standing position induces a downward fluid shift by increasing the hydrostatic pressure gradient, which produces reduced venous return and cardiac output and reduces arterial pressure. This reduction in arterial pressure evokes a baroreflex, and arterial pressure stabilizes [10,11]. Additionally, postural changes stimulate the vestibular receptors, which are also thought to be involved in regulating arterial pressure [1,9,12-14].
The baroreflex sensitivity which is an index of the sensitivity that BP changes lead to heart rate changes reflectivity can reflect the functional status of the cardiovascular autonomic nervous system [15]. Elevations in arterial transmural pressure evoke the discharge of baroreceptors in the carotid sinus and aortic arch, which are sensitive to changes in vessel wall distension. This information is conveyed by branches of the glossopharyngeal and vagus nerves to the rostral ventrolateral medulla (RVLM), resulting in bradycardia and decreased peripheral vascular resistance [16]. The RVLM receives inhibitory inputs from the caudal ventrolateral medulla (CVLM) whose activity is controlled by baroreceptors. Stimulation of baroreceptors by increasing blood pressure results in an increase in CVLM neuronal activity, and a decrease in RVLM activity. This causes sympathetic preganglionic neuronal activity to decrease, thereby lowering blood pressure. The RVLM also contains tonically active presympathetic neurons that monosynaptically innervate the sympathetic preganglionic neurons of the intermediolateral cell column [17]. A direct vestibular projection to the RVLM region has also been reported [18]. A series of studies by Yates and co-workers [19-21] demonstrated that signals coming from peripheral vestibular organs via the vestibular nucleus affect blood pressure, perhaps through the classic pathway of cardiovascular regulation. Electrophysiological and morphological studies have also shown that the RVLM is a key area in the pathway conveying vestibular signals to the gray substance in the spinal cord.
Previous studies in our laboratory have found that sodium nitroprusside (SNP)-induced acute hypotension increases neuronal activities in the medial vestibular nuclei [22], increases expression of c-Fos protein in the medial vestibular nuclei [23], and increases glutamate release in the medial vestibular nuclei [24,25]. These effects are abolished by removing the peripheral vestibular organ. These results indicate that SNP-induced hypotension influences the activity of neurons in the medial vestibular nuclei by increasing afferent signals from peripheral vestibular receptors, and that glutamate, an excitatory amino acid transmitter, in the medial vestibular nuclei is involved in this process. However, the role of the vestibular system compared with the baroreflex in the quantitative regulation of blood pressure is unclear. In addition, the relationship between the vestibular system and the baroreflex in the central nervous system has not been clarified.
Therefore, the effect of bilateral labyrinthectomy and baroreceptor unloading on baroreflex sensitivity was evaluated in this study to investigate the role of the vestibular system compared with the baroreflex in the quantitative regulation of blood pressure. We additionally measured c-Fos protein expression in the RVLM of conscious rats.
METHODS
Animals
Adult male Sprague-Dawley rats (Changchun, China) weighing 220~250 g were used. All animal protocols and procedures described were approved by the Institutional Ethical Committee on Experimental Use of Animals. The animals had free access to food and water. Efforts were made to minimize the number of animals used and suffering.
The rats were divided into four groups to examine baroreflex sensitivity (n=8/group): A sham group, in which both the sinoaortic baroreceptors and vestibular end organs were intact; the bilateral labyrinthectomy (BL) group, in which the sinoaortic baroreceptors were intact, but a bilateral labyrinthectomy was performed; the sinoaortic denervation (SAD) group, in which the sinoaortic baroreceptors were denervated, but the vestibular end organs remained intact; and the SAD+BL group, in which both the sinoaortic baroreceptors and bilateral vestibular end organs were removed. Forty-eight rats were used for an immunohistochemical analysis. Similarly, they were divided into the same four groups (n=12/group). These groups were subdivided into saline infusion as a control and an SNP infusion group (n=6/group).
Labyrinthectomy
A chemical labyrinthectomy was performed as described previously [26]. Briefly, 100 µl of sodium arsanilate (100 mg/ml) was intratympanically injected into the bilateral middle ear of the rats under isoflurane anesthesia (Ilsung Co., Seoul, Korea), which chemically destroyed the membranous labyrinth. The destruction of epithelial cells in peripheral vestibular receptors was confirmed by confocal microscope after rhodamine-palloidin staining. As a control, saline, instead of sodium arsanilate solution, was injected intratympanically in the sham and SAD rats. The labyrinthectomies were performed 48 hours prior to experimentation.
Sinoaortic denervation
The protocol for carotid and aortic sinus denervation was adapted from that described by Wei et al. [27]. Rats were anesthetized with isoflurane. The carotid sinus nerve was sectioned bilaterally following a midventral incision in the neck, and the internal, external, and common carotid arteries were stripped of connective tissue at the level of bifurcation and painted with 10% phenolethanol to denervate the carotid sinus. For aortic arch denervation, the aortic arch nerve was severed bilaterally proximal to its junction with the vagus nerve. In the sham and BL groups, rats received similar cervical incisions leaving nerves, vessels, and baroreceptors intact. After the surgery, animals were breathing spontaneously without significant changes in respiratory rhythm. SAD was performed 24 hours prior to experimentation.
Evaluation of the baroreceptor reflex
A heparinized polyethylene tube was inserted into the femoral artery to record blood pressure in rats under isoflurane anesthesia, and another was placed in the femoral vein for drug infusion. These two polyethylene tubes were connected to the tubes of a cybernation metabolism cage to allow free movement in a conscious state during the experiment. Blood pressure was recorded directly from the femoral artery using a pressure transducer and displayed continuously on a recording system (Powerlab, Sydney, Australia) by a computer. Heart rate (HR) and mean arterial pressure (MAP) were computed from the arterial pressure pulse. Cardiac baroreflexes were evoked by increasing arterial pressures with ramp infusions of phenylephrine (PE; 15 µg/kg/min) and by lowering BP with sodium nitroprusside (SNP; 15 µg/kg/min). Drugs were infused in 3 minutes and BP increased or decreased 30~40 mmHg during this period. Drug order was randomized, and the rats were allowed to recover for 30~40 min before infusion of the next drug was started after the catheter was flushed and reloaded. We took precautions to ensure that the rats rested quietly while the baroreflexes were tested, and that the resting BPs and HRs were stable for 30 min prior to beginning the experiment.
Immunohistochemistry
An immunohistochemical analysis of c-Fos protein expression was performed as described previously [25,28]. Under deep anesthesia with an overdose of sodium pentobarbital, the animals were perfused transcardially with 500 ml of 0.9% NaCl at 4℃ and then perfused with 500 ml of 4% paraformaldehyde dissolved in 0.1 M phosphate buffer (pH 7.4) at 4℃. The brains were dissected and postfixed with paraformaldehyde fixative solution for 4 hours at room temperature. The fixed brains were immersed in 30% sucrose in phosphate-buffered saline (PBS) for 2 days at 4℃ for cryoprotection. The tissue was sectioned to a 20 µm thickness on a freezing microtome (Leica, Nubloch, Germany), incubated for 30 min with 0.3% H2O2, rinsed three times for 5 min with 0.01 M PBS, and then incubated with 1% Triton X-100 for 30 min. After a brief wash, the tissue was incubated for 30 min with 5% bovine serum albumin (PBS+BSA), and then incubated overnight at 4℃ with a rabbit polyclonal anti-c-Fos antibody (1:400; Cell Signaling Technology, Danvers, MA, USA) in PBS+BSA. The following day, tissue sections were incubated with a biotinylated goat anti-rabbit secondary antibody (1:200; Vector Lab., Burlingame, CA, USA) and then with the Elite ABC Kit (Vector Lab.) for 2 hours at 37℃. The bound complex was visualized by incubating the tissue with diaminobenzadine. Sections were then dehydrated, cleared in xylene, and coverslipped with Permount (Fisher Scientific, Pittsburgh, PA, USA). c-Fos positive neurons in the RVLM were counted using a digital image analysis system (Image-Pro; Media Cybernetics, Silver Spring, MD, USA).
Data analysis
Baroreflex sensitivity was estimated by calculating the ratio of changes in HR and MAP before and after PE and SNP infusions (ΔHR/ΔMAP). All data are expressed as mean±standard error. Differences were identified using multivariate analysis of variance. A p<0.05 was considered significant.
RESULTS
After the labyrinthectomy was completed and the animals recovered from anesthesia, they showed severe symptoms of bilateral vestibular loss, such as postural instability and panic behavior. But those acute vestibular symptoms after labyrinthectomy were subsided before recording blood pressure.
Effect of intravenous infusion of PE and SNP on MAP and HR
The baseline MAP and HR values in the sham, BL, SAD, and SAD+BL groups were not different. MAP increased 30~40 mmHg in the four groups (p<0.01) and HR decreased in the sham and BL groups (p<0.01) during intravenous infusion of PE, compared with those at baseline. Nonetheless, the effects were not the same in all groups. HR increased during PE infusion in the SAD and SAD+BL groups (p<0.01) (Table 1). We also compared the SNP-induced decrease in MAP and tachycardic reflex responses. Intravenous infusion of SNP resulted in a significant decrease in MAP in all four groups (p<0.01), and HR increased in the sham and BL groups (p<0.01). HR values were not different from those at baseline in the SAD and SAD+BL groups (Table 2).
Baroreflex sensitivity
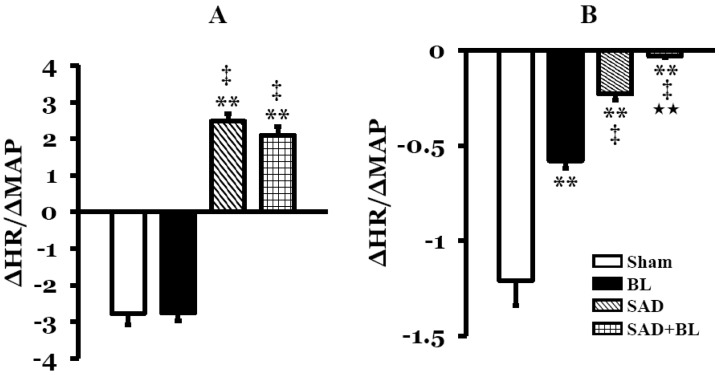
The change in baroreflex sensitivity was measured in the four groups during PE or SNP infusion. Increased baroreflex sensitivity is likely to be an increase in sympathetic activity or a decrease in parasympathetic activity. Baroreflex sensitivity in the sham group response to intravenous infusion of PE was -2.77±0.31, and the values in the BL, SAD, and SAD+BL groups were -2.75±0.22, 2.48±0.21, and 2.09±0.23, respectively. Baroreflex sensitivity in the BL group decreased slightly compared with that in the sham group, but the difference was not significant. Baroreflex sensitivity values in the SAD and SAD+BL groups became positive (p<0.01), which means loss of baroreceptor function. The changes in the SAD and SAD+BL group were due to increased HR during PE infusion. Similarly, baroreflex sensitivity in the SAD and SAD+BL group was significantly different compared with that in the BL group (p<0.01). However, no difference was observed between the SAD and SAD+BL groups (Fig. 1A).
Baroreflex sensitivity values during SNP infusion were all negative. Baroreflex sensitivity in the BL (-0.57±0.04), SAD (-0.18±0.03), and SAD+BL (-0.06±0.04) groups decreased significantly compared with that in the sham group (-1.23±0.14) (p<0.01). Similarly, a significant decrease was observed in the SAD and SAD+BL groups (p<0.01) compared with that in the BL group. A significant difference was also observed between the SAD and SAD+BL groups (p<0.01) (Fig. 1B).
We also observed baroreflex sensitivity in rats that underwent unilateral labyrinthectomy. The results indicate no significant difference compared with the sham group during PE infusion (-2.63±0.19 vs. -2.77±0.31) or SNP infusion (-1.14±0.10 vs. -1.23±0.14).
Effect of intravenous infusion of SNP on c-Fos protein expression in the RVLM
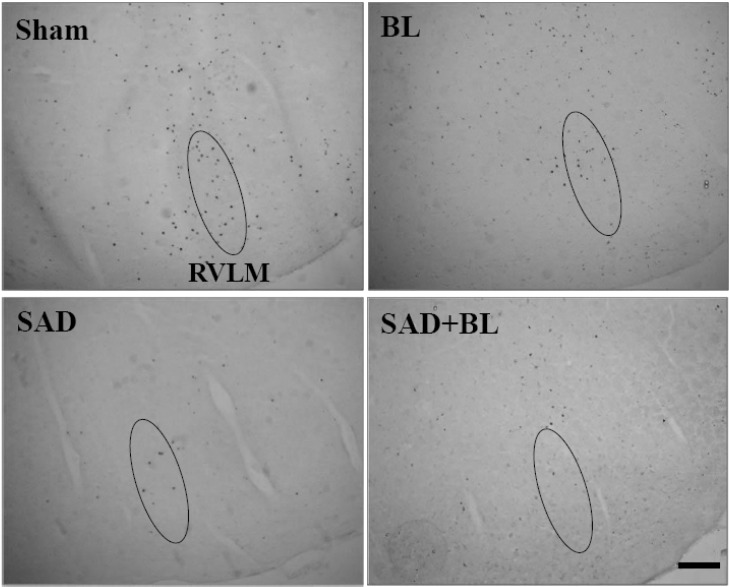
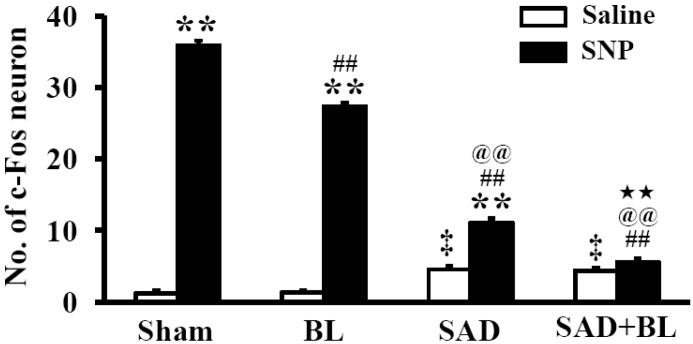
The results of baroreflex sensitivity indicated that the vestibular system may participate in the regulation of acute hypotension. Therefore, expression of the c-Fos protein was observed in the RVLM following acute hypotension induced by SNP. The results of a preliminary experiment suggested that c-Fos protein expression in the RVLM at 30 min (9.54±0.43), 60 min (23.71±0.32), 90 min (35.89±0.65), and 120 min (21.41±0.70) was different after acute hypotension. c-Fos protein expression peaked 90 min after SNP infusion. Consequently, the experiment was carried out during this time period (Fig. 2).
In the sham group, intravenous administration of saline at the same volume and rate as SNP infused in the experimental groups did not induce significant expression of the c-Fos protein in the RVLM (1.22±0.32). Significantly greater expression of the c-Fos protein was observed in the sham group after SNP infusion (35.89±0.65) (p<0.01). Similarly, little c-Fos protein was expressed in the RVLM by infusion of saline in the BL group; however, significant expression of the c-Fos protein was observed following infusion of SNP (27.33±0.53 vs. 1.33±0.29, p<0.01). Notably, c-Fos protein expression in the SAD and SAD+BL groups following infusion of saline increased clearly compared with that in the sham group infused with saline (4.56±0.38 vs. 1.22±0.32, p<0.05; 4.37±0.33 vs. 1.22±0.32, p<0.01). c-Fos protein expression in the SAD group following infusion of SNP increased significantly compared with infusing saline (11.12±0.59 vs. 4.56±0.38, p<0.01). However, this phenomena was not observed in the SAD+BL group, and no significant difference was observed between the SNP and saline infusions (5.56±0.44 vs. 4.37±0.33). A comparison within groups infused with SNP indicated that c-Fos protein expression in the BL, SAD, and SAD+BL groups was greatly reduced compared to that in the sham group (p<0.01). Furthermore, the SAD and SAD+BL groups decreased clearly compared to the BL group (p<0.01), and a significant difference was observed between the SAD and SAD+BL groups (p<0.05) (Fig. 3).
DISCUSSION
In the present study, we found that: (1) BL, SAD, and SAD+BL did not affect resting MAP and HR. (2) SAD caused a loss of the baroreflex for regulating PE-induced hypertension. (3) BL, SAD, and SAD+BL resulted in decreased baroreflex sensitivity during intravenous infusion of SNP. (4) SNP-induced acute hypotension increased c-Fos protein expression in the RVLM of the sham group; however, BL, SAD, and SAD+BL attenuated c-Fos protein expression. Among them, c-Fos protein expression clearly decreased in the SAD+BL group.
The vestibular system modulates blood pressure through the sympathetic nervous system [29]. However, patients with bilateral vestibular loss complain of orthostatic hypotension [30], and animals with BL have reduced arterial pressure when body posture changes from a lying to a standing position [9]. These reports indicate that the vestibular system plays an important role regulating blood pressure during postural change. Moreover, we reported that acute hypotension induced by hemorrhage or infusion of SNP increases neuronal activity, c-Fos protein expression, and pERK expression in the medial vestibular nuclei, but that these effects are abolished following BL [22,23,31]. These studies indicate that the peripheral vestibular receptors respond to decreases in blood pressure.
The effect of SAD on the conscious primate is controversial; some studies have reported that SAD increases blood pressure [32,33] but others SAD has no change in blood pressure [34-36]. It is considered that major differences between the studies are the denervation procedure and the conditions under which blood pressure was recorded. Theoretically, removal of the baroreceptor input should result in a hypertension since the inhibitory inputs from the CVLM whose activity is controlled by baroreceptors are abolished [17]. However, when the influence of baroreceptors is removed, cardiopulmonary receptors may assume an important role [35]. In the present study, SAD did not change resting MAP or HR, which could be due to denervation procedure and stable recording conditions. PE induces a reflex bradycardia when the baroreceptors are intact, but not when they are denervated. Baroreflex sensitivity was higher during PE-induced hypertension than during SNP-induced hypotension in sham animals. BL attenuated baroreflex sensitivity during intravenous infusion of SNP, but did not affect sensitivity during intravenous infusion of PE. SAD and SAD+BL decreased sinoaortic baroreflex function during PE infusion, but did not completely eliminate the baroreflex effect during SNP-induced hypotension. The vestibular system still plays a significant role modulating blood pressure during SNP-induced hypotension in baroreceptor-denervated animals but has no effect in PE-induced hypertension. Vestibular-elicited alterations in sympathetic nerve activity can produce changes in blood pressure and blood flow to specific vascular beds [2,37]. The vestibulo-sympathetic reflex pathways are independent from baroreflex pathways [6,38]. Thus, the baroreflex sensitivity results in the baroreceptor-denervated animals might be due to activation of pathways between the vestibular system and other areas of the central nervous system participating in cardiovascular modulation.
The immediate early gene protein c-Fos is useful for mapping functional activity of the brain [39,40]. Several studies have demonstrated that hypotension elicits c-Fos protein expression in several forebrain and brain stem nuclei that affect cardiovascular function [23,41]. Previous studies in our laboratory have found that SNP-induced acute hypotension increases c-Fos and Erk protein expression in the medial vestibular nuclei [23,25,31,42].
SNP decreases blood pressure by causing the release of nitric oxide [43]. SNP-induced hypotension increases c-Fos protein expression in the supraoptic nucleus, paraventricular nucleus, RVLM, solitary tract nucleus, and vestibular nucleus [44,45]. However, PE-induced hypertension decreases c-Fos protein expression in the RVLM [44], suggesting that c-Fos protein expression in the RVLM caused by SNP-induced hypotension is a baroreflex pathway.
The RVLM, which contains many barosensitive neurons, is mostly activated by hypotension and inhibited by hypertension [46], and it is a mediator of peripheral baroreceptors and vasomotor centers [47]. Anatomical evidence shows the existence of a direct projection from vestibular nuclei to the RVLM region [40,48]. Functional convergence from the vestibular and baroreceptor afferents to the same nuclei in the RVLM further suggests that this is the central point of interaction between the vestibular system and the baroreflex [38]. In the present study, a significant increase in c-Fos protein expression in the RVLM was observed in the sham group after administration of SNP. However, c-Fos protein expression decreased significantly in the BL, SAD, and SAD+BL groups, and particularly in the SAD+BL group. These results indicate that the vestibular signals caused by SNP-induced hypotension are probably mediated by vestibular nuclei in the RVLM besides the baroreceptor reflex. That is to say, vestibulo-cardiovascular reflex pathways from the peripheral vestibular organ-vestibular nuclei-RVLM exist in the central nervous system independent of the baroreflex. Studies have shown that vestibulo-sympathetic reflexes are patterned according to both the anatomical location and innervation target of a particular sympathetic nerve and could lead to distinct changes in local blood flow [2,5,29]. Neurons in the RVLM of cats, which are components of the baroreceptor reflex circuitry, also mediate vestibulo-sympathetic responses [1,49]; the processing of both baroreceptor and vestibular signals by these cells likely explains why the two reflex mechanisms have additive effects on cardiovascular control [5,50]. Neurons in the RVLM that regulate blood pressure have fewer sensory inputs available after damage to the vestibular nuclei.
In summary, we observed the effects of labyrinthectomy and SAD on baroreflex sensitivity and c-Fos protein expression in the RVLM following acute hypotension or hypertension in freely moving conscious animals. Based on our previous studies, we found that the vestibular system participates in regulating blood pressure, particularly during acute hypotension. Furthermore, both the baroreflex and vestibular system cooperate to maintain arterial pressure during SNP-induced hypotension. Nevertheless, the exact mechanism of blood pressure regulation by the vestibular system via the RVLM requires further research.


 XML Download
XML Download