

 PDF
PDF ePub
ePub Citation
Citation Print
Print


ABBREVIATIONS
TEA
tetraethylammonium
4-AP
4-aminopyridine
OXT
oxytocin
RMP
resting membrane potential
KCa channel
Ca2+-activated K+ channel
KV channel
voltage-activated K+ channel
K2P channels
two-pore domain weak inwardly rectifying K+ channel
TASK
two-pore domain weak inwardly rectifying K+ channel (TWIK)-related acid-sensing K+ channels
TREK
stretch-dependent K2P channels
KRB
Krebs-Ringer bicarbonate
MES
2-N-morpholinoethanesulphonic acid
pHo
extracellular pH
TTX
tetrodotoxin
ATR
atropine
NIOK
non-inactivating outward K+ current
VDCCL
voltage-dependent L-type Ca2+ channel
INTRODUCTION
Regulation of myometrial contractility, which maintains a relaxed state during pregnancy and then shows highly organized contraction at labor and delivery, is an important process for normal delivery. Myometrial dysfunction during pregnancy leads to premature delivery, which is a major risk factor for newborn deaths [1]. Risk factors for preterm delivery include working during pregnancy, smoking, inadequate maternal weight gain, illicit drug use, and genetic factors [2]. Unfortunately, there is no effective means of preventing preterm delivery. Atociban, an oxytocin (OXT) receptor antagonist, is still unsatisfactory as a major therapeutic medicine [2].
Myometrial contractility is controlled by a complex interplay among many humoral factors and hormones [3]. First, plasma pH rises during pregnancy [4]; however, normal labor at term is associated with the development of maternal acidemia [5,6]. One of the explanations for the acidemia is ischemia [7] due to occlusion of blood vessels caused by strong myometrial contractions [8]. Second, complex hormonal changes occur during pregnancy. The pregnant myometrium is quiescent mainly due to increased progesterone [9] until the beginning of highly organized contractions at labor [10]. Additionally, the increase in estrogen from mid-gestation to birth is to myometrial contractility [11]. Therefore, extracellular pH and sex hormones are important factors regulating myometrial contractility during pregnancy [12-14]. However, the regulatory mechanisms of myometrial contractility at the level of the ion channel under pH changes are still poorly understood.
Myometrial contractile responses in mammals are generated by changes in ionic conductance [15]. K+ channels play an important role regulating the resting membrane potential (RMP) of smooth muscle and their excitability; activation of K+ channels decreases membrane excitability via the outward K+ current. Several types of K+ channels that are activated by diverse intracellular factors such as Ca2+ (KCa channels) and voltage (KV channels), have been reported in smooth muscle cells including myometrium [16-19]. In particular, the BK channel, one of the KCa channels, is active at rest thereby maintaining myometrial quiescence. However, changes in its characteristic such as loss of Ca2+ and voltage dependence occur at the onset of labor [16,18].
Among several K+ channels, the most recently identified K+ channel member is the two-pore domain K+ (K2P) channel [20,21], which is thought to contribute to the RMP. K2P channels can be divided into six subfamilies [21]. One of them, TASK (TWIK(two-pore domain weak inwardly rectifying K+ channel)-related acid-sensing K+ channels) channels have been identified in both electrically excitable and non-excitable tissues [21]. TASK-2 channels can be inhibited pharmacologically by several inhibitors [21], and they are exquisitely sensitive to perturbations in extracellular pH [22]. In addition to TASK channels, TREK-1 channels are also important for regulating membrane excitability of the myometrium [23] and are affected by various factors such as extracellular pH and membrane stretch (stretch activated channels, TREK-1) [21,22].
Since remarkable changes such as pH, excitability via activation of ion channels including K2P channels in pregnancy were reported, we tried to elucidate involvement of TASK-2 channels in uterine circular muscle of mouse.
METHODS
Tissue preparation for isometric contraction
Female non-pregnant ICR (10~12 weeks old) and pregnant ICR mice (19 days) were anaesthetized with isoflurane and sacrificed by cervical dislocation. All experiments were performed in accordance with the guidelines for animal care and use approved by Chungbuk National University. The uteri were cut open from the neck to the end of the uterine horns. Tissues were rinsed in Krebs-Ringer bicarbonate (KRB) solution and then pinned down on a Sylgard plate to maintain their original shape and length. Connective tissue from the uteri was removed and the muscle layers were isolated from the endometrium. For the measurement of mechanical contractions, strips (0.2×0.7 cm) of circular muscle were mounted in an organ bath (25 ml) of an isometric contractile measuring system [24]. In this system, one end of the tissue was tied tightly to a fixed holder and the other side was linked by a hook type holder to a force transducer (Harvard Apparatus, Holliston, MA, USA). The force transducer was connected to a PowerLab-Data Acquisition System and Charter v5.5 software (ADInstruments, Boulder, CO, USA) with an IBM compatible computer to measure isometric contractions. Each strip was stretched gradually for 1.5~2 hours to achieve resting tension, and then the contractile response was tested by applying a high K+ (50 mM) solution to the strip, which was repeated three or four times until the responses reached a maximum level.
Immunohistochemical labeling of K2P channel (TASK-2)
To visualize cell expressing immunoreactivity of TASK-2 channels, both non-pregnant and pregnant uterus were used. For immunohistochemistry, murine uterus were cut and pinned to rectangular shape (2×3 cm, width and length) in KBR solution and tissues were fixed in 4% paraformaldehyde in 0.1 M sodium phosphate buffer for 24 hours at room temperature. Two circular sections, 3 cm in length and 3 mm in thickness, were taken from each fixed tissue. Sections were cut at 4 µm thickness with a microtome from paraffin-embedded tissue blocks and mounted on positively charged slides (Superfrost Plus; VWR Internatinal, West Chester, PA, USA). Simultaneous deparaffinization and antigen retrieval prior to immunostaining were accomplished on an automated PT module (Lab Vision, Fremont, CA, USA). Immunohistochemistry was conducting using an automated immunostainer (Autostainer 360; Lab Vision) accounting to the manufacturer's protocol. Peroxidase staining was carried out using UltraVision LPDetection System HRP Polymer & DAB Plus Chromogen (Thermo Fisher Scientific, Fremont, CA, USA). Briefly the sections were incubated in Hydrogen Peroxide Block for 10 minutes to reduce nonspecific background staining due to endogenous peroxidase. After washing in phosphate buffered saline (PBS) plus Tween 20 (20×) (ScyTek laboratories, Logan, Utah, USA), they were incubated with Ultra V Block for 5 min at room temperature to block nonspecific binding. The sections were then incubated with TASK-2 primary antibody (SantaCruze, USA) at a dilution of 1:200 (room temperature). After washing in PBS, they were incubated at room temperature with primary antibody enhancer, followed by washes in PBS and incubation with the HRP polymer for 15 min at room temperature. Sections were then washed in PBS and followed by staining with DAB plus chromogen and substrate. Counterstaining was performed with hematoxylin. Negative controls were performed by omitting the primary antibodies or by substitution them with a non-immune serum in orderto check the specificity of the immunostaining. All sections for histologic and immunohistochemical analysis were examined using a microscope (BX50; Olympus Corporation) and images were captured with an attached camera (ProgRes C14; Jenoptik, Jena, Germany) operated with CapturePro software (Jenoptik) [25,26].
Tissue preparation for sharp electrode recording
The endometrium removed tissue was pinned out on a silicon rubber plate fixed at the bottom of the organ bath, with the circular muscle uppermost. The tissue was superfused with warmed (36℃) and oxygenated Krebs solution, at a constant flow rate of about 2 ml/min. Conventional microelectrode technique was used to record intracellular electrical responses of uterine circular muscle. Briefly, glass capillary microelectrodes (outer diameter, 1.2 mm, inner diameter 0.6 mm; Hilgenberg, Germany) filled with 0.5 M KCl, with the tip resistances ranging between 50~80 MΩ, were inserted to cells. Electrical responses recorded were amplified through a high input impedance amplifier (Axoclamp-2B, Axon Instruments, Foster City, Calif., USA), and were also displayed on a cathode-ray oscilloscope (SS-7602, Iwatsu, Osaka, Japan). The responses were also stored on a personal computer for later analysis of the data.
Solution and drugs
KRB solution (CO2/bicarbonate-buffered Tyrode) contained (in mM) NaCl 122, KCl 4.7, MgCl2 1, CaCl2 2, NaHCO3 15, KH2PO4 0.93, and glucose 11 (pH 7.3~7.4, bubbled with 5% CO2/95% O2). The membrane impermeable pH buffer, 2-N-morpholinoethanesulphonic acid (MES, Sigma Chem Co., St Louis, MO, USA) was used to adjust the KRB solution to pH 6.4 when necessary. Equimolar concentrations of external Na+ were replaced by K+ for 50 mM K+ solution. The external solution was changed to the next solution already pre-incubated (bubbled with 5% CO2/95% O2, 36℃) in water bath before application. All drugs used in this study were purchased from Sigma except for antibodies against these proteins such as TASK-2 channels (SantaCruze, USA), Na+/K+-ATPase (SantaCruze, USA) (Thermo Scientific, USA).
Statistics
Data were expressed as means±standard error of the mean (means±SEM). The Student's t-test (paired and unpaired) was used wherever appropriate to evaluate differences in data. Wilcoxon rank-sum test and Mann-Whitney test was also used. p values less than 0.05 were regarded as statistically significant.
RESULTS
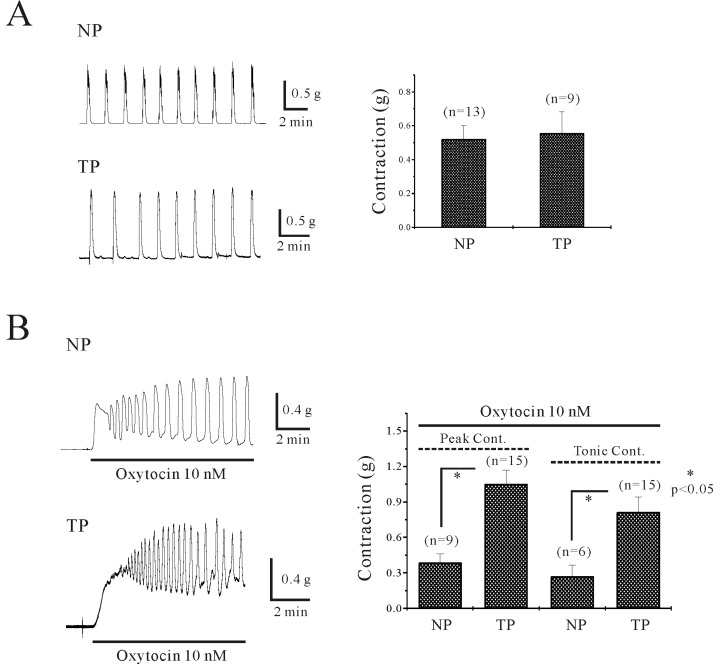
Characterization of oxitocin (OXT)-induced contraction of uterine circular muscle
Uterine circular muscle shows spontaneous phasic contraction in both non-pregnant (0.5±0.08 g, 0.9±0.13 cycles/min; n=13 and 13) and pregnant myometrium (0.6±0.13 g, 1.4±0.17 cycles/min; n=9 and 9) (Fig. 1A). However spontaneous contraction was abolished by application of step wise stretch (see methods). OXT produced tonic contractions superimposed with phasic contractions in both tissues. Initial peak contractions induced by OXT were 0.4±0.08 g (nonregnant, n=9) and 1.1±0.12 g (pregnant, n=15), respectively. Tonic contractions were 0.3±0.10 g (non-pregnant, n=6) and 0.8±0.13 g (pregnant, n=15) (Fig. 1B). Phasic and tonic contractions induced by OXT were significantly different in non-pregnant and pregnant myometrial tissues (p<0.05).
Effect of quinidine on spontaneous electrical activity of uterine circular muscle
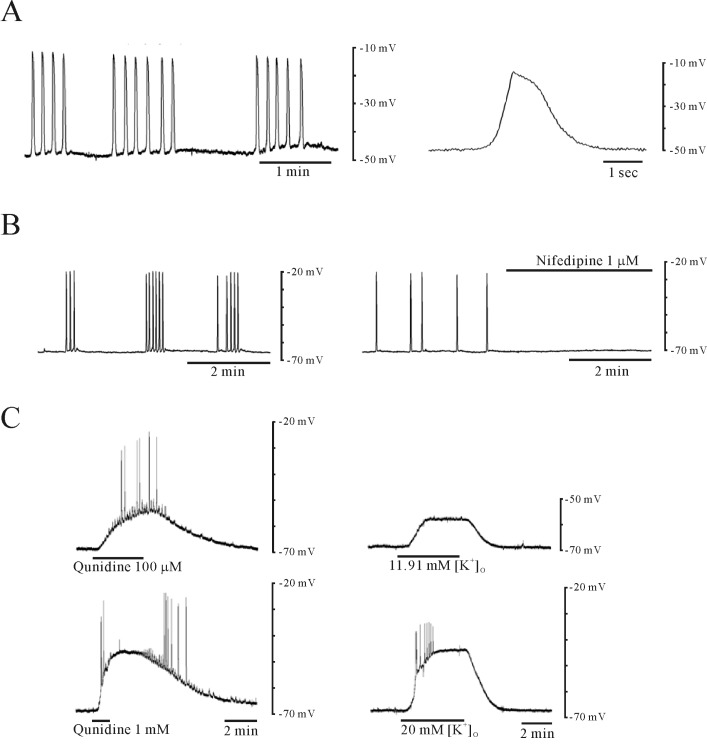
Quinidine produced concentration-dependent depolarization in circular muscle under sharp electrode recording (Fig. 2C). Circular muscle showed RMP of -69±3.5 mV (n=5) and spontaneous active action potential which was sensitive to nifedipine (1 µM) (Fig. 2A and 2B). Application of quinidine (100 µM) produced depolarization of 15±2.3 mV (n=5). Quinidine (1 mM) produced depolarization over 20 mV (Fig. 2C) however quinidine (10 µM) produced depolarization less than 10 mV (data not shown). In right panel of Fig. 2C, high K+ depolarized the membrane, in a concentration-dependent manner, with generation of spike potentials.
Effect of quinidine and extracellular acidosis on isometric contraction of uterine circular muscle
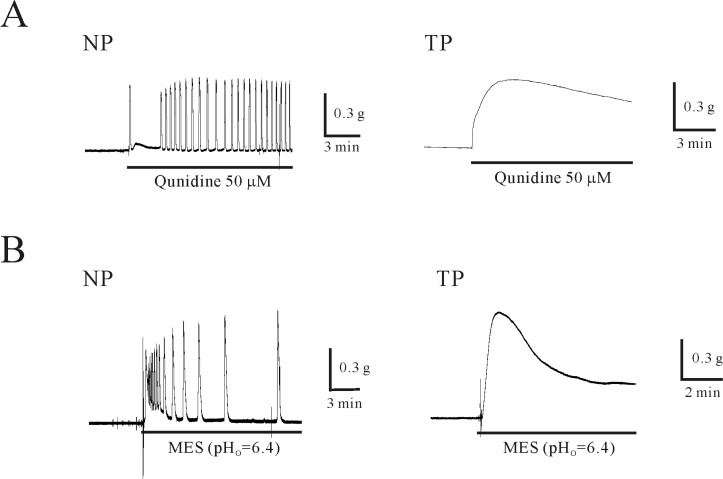
The effect of quinidine and extracellular acidosis, an inhibitor of the K2P channel, on uterine circular muscle was studied [21]. As shown in Fig. 3A, quinidine (50 µM) provoked contractions in non-pregnant and pregnant uterine circular muscles. Quinidine-induced contractions began to merge to tonic contractions, and this tendency was stronger in pregnant uterine circular muscle. In two cases, quinidine (50 µM) slightly increased the magnitude of tonic contractions in non-pregnant uterine circular muscle (0.07±0.04 g). However, quinidine-induced tonic contraction was significantly increased in pregnant uterine circular muscle (0.3±0.07 g, n=8; p<0.05).
Because quinidine is a potent inhibitor of TWIK channels (TASK; IC50=22 µM for TASK-2 in human kidney) [27], we also studied the effects of other TASK channel inhibitors such as extracellular acidosis. Extracellular pH was changed to acidic condition (pHo=6.4) and regulatory effect of extracellular acidosis on uterine circular muscle was studied. As shown in Fig. 3B, extracellular acidosis produced phasic contractions (0.6±0.10 g) in non-pregnant uterine circular muscle (n=9). In pregnant uterine circular muscle, extracellular acidosis produced peak (1.0±0.09) and phasic contractions (0.9±0.22 g) (n=11 and 3, respectively). The tonic contraction of non-pregnant and pregnant uterine circular muscle were 0.02±0.005 and 0.2±0.04 g (n=5 and 10; p<0.05).
Effect of tetraethylammonium (TEA) and 4-aminopyridine (4-AP) which inhibits of Ca2+-activated K+ (KCa) channel and voltage-dependent K+ (KV) channel on uterine circular muscle of mouse
Since Ca2+- (KCa) and voltage-activated K+ (KV) channel in smooth muscle exist and its activation attenuate excitability including alteration of its characteristic in labor [19,28,29], we also designed to study role of TASK-2 inhibitors in the presence of TEA and 4-AP which block KCa channel and KV channel in uterine circular muscle.
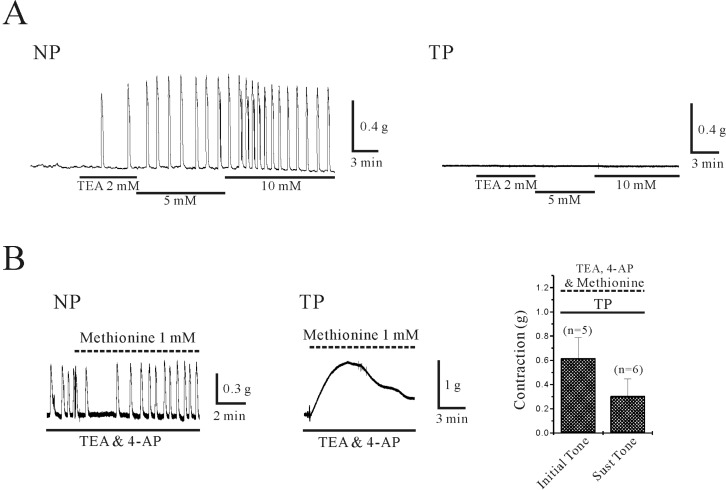
In non-pregnant and pregnant uterine circular muscles, effect of TEA and 4-AP were studied. As shown in Fig. 4A, TEA produced concentration dependent phasic contractions in non-pregnant myometrium. TEA (2, 5 and 10 mM) produced contractions of 0.7±0.09, 0.6±0.10 and 0.7±0.09 g (n=7, 8 and 9) with a frequency of 0.5±0.10, 0.7±0.19 and 0.7±0.15 cycles/min (n=7, 8 and 9). However, term pregnant myometrium did not show any contractile effect by TEA (1~10 mM) (n=10). 4-AP also produced contraction in non-pregnant uterine circular muscle. 4-AP (5 mM) produced phasic and tonic contraction of 0.8±0.35 g and 0.12±0.08 g (n=2 and 4; data not shown). However, 4-AP (5 mM) produced tonic contraction in pregnant uterine circular muscle (0.8±0.16 g, n=7).
As shown in Fig. 4B, effect of L-methionine known to inhibit stretch-activated channel on myometrial circular muscle was also studied in the presence of TEA and 4-AP. In the presence of TEA (10 mM) and 4-AP (5 mM), L-methionine (1 mM) did not show significant effect in non-pregnant uterine circular muscle (n=6). However, L-methionine (1 mM) in the presence of TEA and 4-AP produced transient contraction (0.6±0.17; n=5), which decayed to a low level (0.3±0.14g; n=6) within 10 min.
Regulation of murine pregnant myometrial contractility by TASK-2 channel inhibitors
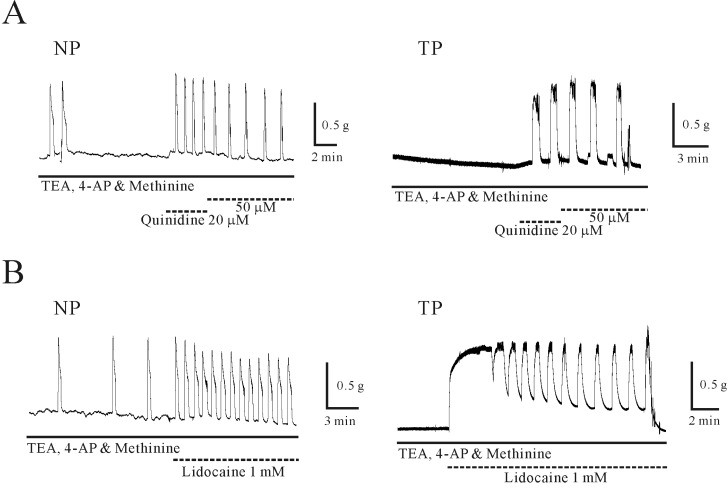
To rule out the involvement of other K+ channels except the TASK-2 channels in the response to the TASK inhibitors [27], the TASK channels were inhibited with quinidine and lidocaine in the presence of TEA, 4-AP and L-methionine in uterine circular muscle. As shown in Fig. 5A and 5B, quinidine and lidocaine provoked contractions in the presence of TEA, 4-AP and L-methionine.
In the presence of TEA, 4-AP and L-methionine, quinidine (50 µM) produced phasic contractions of 0.5±0.15 g in non-pregnant circular muscle (n=3; Fig. 5A). In pregnant tissue, quinidine (50 µM) produced phasic and tonic contraction of 0.4±0.21 g and 0.1±0.11 g (n=4 and 2). Lidocaine (1 mM) also produced contraction in the presence of TEA, 4-AP and L-methionine (Fig. 5B). In non-pregnant circular muscle, phasic contraction (0.6±0.28 g) was produced by lidocaine (n=4). Meanwhile, lidocaine produced peak, phasic and tonic contractions in pregnant circular muscle (1.0±0.26 g, 1.0±0.26 g and 0.2±0.07 g; n=3, 5 and 3, respectively).
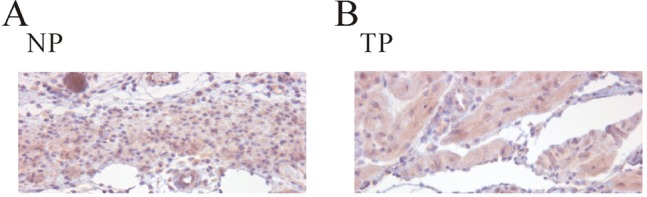
Identification of TASK-2 by immunohistochemistry in uterine circular muscle of mouse
Finally, we confirmed expression of TASK-2 channels by immunohistochemistry. TASK-2 expression was observed in myometrial circular smooth muscle of both non-pregnant and pregnant mouse. As shown in Fig. 6, its expression in non-pregnant mouse is sparse however expression of TASK-2 was increased evenly to whole cell membrane in pregnancy.
DISCUSSION
K2P channels are regulated by various factors such as extracellular pH [22] and membrane stretch [30]. The pharmacological characteristics of TASK-2 channels, one of the K2P channels, are inhibited by quinidine, lidocaine, and extracellular acidosis [21]. Our experiment showed similar results and also showed strong TASK-2 channels expression by immunohistochemistry during pregnancy. This is the first study proposing extracellular pH-sensitive K+ conductance as key regulators of pregnant myometrium.
Contractions of uterine muscle in mammals are generated from changes in ionic conductance [15,31]. In this study, we studied effect of quinidine, lidocaine, and extracellular acidosis on isometric contraction of uterine circular muscle to evaluate functional expression of TASK-2 channels in mouse. As shown in Fig. 3A, quinidine produced contraction in both non-pregnant and pregnant uterine circular muscles. In addition, contractile effect of quinidine was enhanced in pregnant uterine circular muscle compared to non-pregnant tissue. Similar to the effect of quinidine, enhanced contractile effect of extracellular acidosis was also observed in pregnant uterine circular muscle (Fig. 3B). In Fig. 5, we also studied effect TASK-2 channels inhibitors in the presence of other K+ channels blockers. K+ channels such as Ca2+- and voltage-activated K+ channels (KCa and KV) are ubiquitous in smooth muscle cells and play an important role regulating the RMP of smooth muscle and its excitability. K2P channels are also thought to contribute to the RMP [31]. As activation of K+ channels suppresses membrane excitability in vessels [28, 29] as well as the myometrium at labor [16,18,32,33], we examined the effect of TASK-2 inhibitors on contractility of uterine circular muscle in the presence of TEA and 4-AP. In accordance with previous reports showing that most K2P channels are insensitive to TEA and 4-AP [21], application of TEA and/or 4-AP produced no contractions except for weak transient contractions in some pregnant muscle cells (Fig. 4) [32]. As shown in Fig. 5, TASK-2 channels inhibitors such as quinidine, and lidocaine produced robust contractions even in the presence of TEA, 4-AP, and/or L-methionine. Even data not shown, these contractile effect of TASK-2 channels inhibitors on uterine contraction was not affected by nerve blockers such as nerve blocker cocktail (0.4 µM tetrodotoxin (TTX), 1 µM guanethidine, and 1 µM atropine (ATR)) [24]. This result strongly suggests that TASK-2 channels might play a central role in the regulation of uterine circular muscle contractility.
As shown in Fig. 4, TEA and 4-AP produced phasic contraction in non-pregnant circular muscle but it did not show strong activity in pregnant circular muscle. To date, change of characteristics of KCa channel via loss of Ca2+ and voltage dependency at the onset of labor [16,18]. Therefore, active KCa channels and/or KV channels at resting state particularly maintain uterine quiescence in non-pregnant uterus. Meanwhile, other K+ channel activity such as TASK channels, TREK-1 channels (stretch activated channels) which are affected by various factors such as extracellular pH and membrane stretch [21,22] might be important for regulating membrane excitability of the uterine muscle [23]. Although data not shown in here, we already observed non-inactivating outward K+ current (NIOK) insensitive to TEA and 4-AP may be responsible for the quiescence of pregnant murine myometrium in single cell level. The characteristics of NIOK coincided with two-pore domain acid-sensing K+ channels (TASK-2) since NIOK was inhibited by TASK-2 inhibitors under K+ channel blockers. Therefore, diverse channels might be related to regulation of myometrial excitability [12,16,18]. Further evidenced will be revealed in the future.
Uterine smooth muscle is electrically excitable and produces spontaneous contraction which is regulated by ion channels [34,35]. It shows resting membrane potential (RMP) of -69 mV (n=5) [12] and nifedipine (1 µM) inhibited spontaneous action potential in circular smooth muscle (Fig. 2A and 2B). Quinidine (100 µM) which is known to inhibit TASK-2 channels produced 15 mV depolarization (n=5, Fig. 2C). Our mechanical study using uterine circular muscle also revealed that quinidine produced phasic contraction and contractile effect was increased with tonic contraction in pregnant uterine circular muscle (Fig. 3A and 5A). Therefore, membrane depolarization by inhibition of TASK-2 channels might responsible for quinidine-induced contraction in uterine circular muscle of mouse.
Plasma pH tends to be altered during pregnancy and at labor [4-6]. Our data indicate myometrial relaxation through activation of extracellular pH-sensitive TASK-2 channels during pregnancy. In previous reports, decreased pHo abolished phasic contractions and increased pHo induced tonic contractions in rats and humans [36,37]. Increased pHo produces relaxation by inhibiting voltage-dependent L-type Ca2+ channels (VDCCL) [14]. In this study, we found decreased pHo enhanced myometrial contraction of mouse. However, the effect of extracellular pH is difficult to distinguish from intracellular pH as changes in extracellular and intracellular pH interact with each other [14,38].
Uterine circular muscle showed spontaneous contractions in non-pregnant (0.5±0.08 g, 0.9±0.13 cycles/min; n=13 and 13) and pregnant myometrium (0.6±0.13 g, 1.4±0.17 cycles/min; n=9 and 9) (Fig. 1A); however, it was abolished with stretch (see Methods) in both tissues (Fig. 4A). This phenomenon can be explained by TREK-1. The increased expression and function of stretch-dependent K2P channels (TREK-1) has been reported in the myometrium during pregnancy [12,23,39]. In addition to the role of stretch activated K2P channels, this is the first report relating TASK-2 channels to a functional mechanism of relaxation during pregnancy. We showed strong contractions using TASK-2 channels inhibitors in the presence of TREK-1 channels inhibitors (Fig. 5). Therefore, the increased expression of TASK-2 channels might also be a reasonable explanation for uterine accommodation of the growing fetus during pregnancy rather than the increase in fetus-induced passive response to stretch-mediated channels.
 XML Download
XML Download