

 PDF
PDF ePub
ePub Citation
Citation Print
Print


ABBREVIATIONS
EGCG
(-)-epigallocatechin-3-gallate
TT/D
testicular torsion and detorsion
TT
testicular torsion
IAP
inhibitor of apoptosis
BIR
baculovirus repeat
RT-PCR
reverse transcription-polymerase chain reaction
TBS
testicular biopsy score
STD
seminiferous tubule diameter
GECT
germinal epithelial cell thickness
i.p.
intraperitoneal (i.p.)
TUNEL
terminal transferase-mediated dUTP-biotin nick end-labeling
H&E
hematoxylin and eosin
ANOVA
a one-way analysis of variance
GCA
germ cell apoptosis (GCA)
INTRODUCTION
(-)-Epigallocatechin-3-gallate (EGCG) is the major polyphenol extracted from green tea. Several studies have established its numerous properties, including antioxidant, anti-inflammatory, anti-tumorigenic and immunosuppressive effects [1]. The anti-proliferative property of EGCG has made it a plausible chemotherapeutic agent for many cancer models both in vivo and in vitro [2-4]. This growth inhibitory effect has been shown to affect only cancerous cells but not their normal counterparts [5]. In a previous study, the anti-apoptotic effect of EGCG was demonstrated in a non-cancer condition, testicular torsion (TT) [6]. TT is a common urologic emergency that requires immediate surgical intervention to save the testicles. However, delayed action may increase the risk of damage to spermatogenesis, which could result in testicular atrophy and future infertility [7,8]. Survivin, an inhibitor of apoptosis (IAP), has been identified as one of the pro-survival molecules affected by the EGCG protective action [6]. It has a unique dual function as both a regulator of cell division and an IAP [9].
Survivin gene transcription gives rise to three splice variants, which have been identified in humans [10], mice [11] and chicken [12] with high sequence homology and which are expressed in most tissues of the body, including testicular tissue. The human (142) and mouse survivin (140) share approximately 83.1% homology. Both proteins contain a baculovirus repeat (BIR) domain and an α-helical coiled-coil domain and lack a COOH-terminus RING finger domain. The other survivin variant is human survivin-2B and its mouse homolog is survivin 121; both lack the coiled-coil domain but still retain their IAP function. Human survivin variant ΔEx3 and its mouse counterpart survivin 40 are both missing exon 3 and have a markedly reduced antiapoptotic activity. Interestingly, little is known regarding the differential function or differential expression of the survivin splice variants or how they might interact with other proteins or themselves.
The aim of this study was to determine which survivin splice variant is associated with the protective role of EGCG in TT/D. Thus, we evaluated the expression of the three survivin splice variants (140, 121 and 40) using semi-quantitative RT-PCR. Interestingly, the expression of survivin variant 121 was not previously detected in mouse testes [13] or sciatic nerve [14]. However, we attempted to measure its expression in this study to exclude differences in the mouse strains, tissue types and experimental conditions. In support of TT/D-induced apoptosis, we also assessed the expression of some apoptosis genes and monitored the level of DNA fragmentation. To further explore the protective role of EGCG on spermatogenesis during TT, we performed a morphometric analysis of the seminiferous tubules to measure the testicular biopsy score (TBS), seminiferous tubule diameter (STD) and germinal epithelial cell thickness (GECT).
METHODS
Animals
In this study, 18 adult male MF1 albino mice (30~40 g) were housed in a controlled environment in a 12 h light/12 h dark cycle and provided with standard rodent chow and water ad libitum. Animal care was in compliance with the guidelines of experimental animals recommended by Kuwait University for animal welfare and care. All animal experiments followed a protocol approved by the ethics committee on animal research at Kuwait University. The animals were randomly divided into three groups (n=6 each): group I=sham; group II=TT/D+vehicle and group III=TT/D+EGCG.
Surgical procedure
All operations were performed under sterile conditions with the aid of a surgical microscope, as described previously [6]. The animals were anesthetized with an intraperitoneal (i.p.) injection of a mixture of ketamine (62.5 mg/kg) and xylazine (3.2 mg/kg). The skin of the surgical site was shaved and cleaned with 70% alcohol and betadine. In the sham group, a left-sided ilioinguinal incision was made, and the left testis was brought out through this opening and exposed for 60 min before it was positioned back into its scrotal sac. Then, the incision was sutured. The TT/D+vehicle animals were subjected to 60 min of unilateral ischemic injury to the left testes by compressing the testicular artery with a jeweler's microforceps (Fine Science Tools Inc., North Vancouver, BC, Canada) to completely cut off the blood supply to the testes. These animals were i.p. injected with 250 µl of saline 30 min after the beginning of the ischemic injury. The mice in the TT/D+EGCG group were subjected to a unilateral ischemic injury of the left testis for 60 min and i.p. injected with 50 mg/kg EGCG (dissolved in normal saline) 30 min after the onset of injury. The microforceps were released at the end of the 60 min, the testes were placed back into the scrotal sacs, and the wounds and skin incisions were sutured. The testes from all three animal groups were covered with saline-wet gauze to prevent any damage that may result from dryness during the entire 60 min of ischemia. Subsequently, the testes from all animal groups were allowed 4 h of reperfusion before harvesting. The left and right testes from each animal were dissected out, and each testis was cut into two halves for further analyses. The contralateral right testis served as a positive internal control.
RNA isolation and RT-PCR
Total RNA was purified from testicular tissue using the Trizol reagent (Life Technologies Inc., Gaithersburg, MD, USA) following the manufacturer's protocol. Semi-quantitative RT-PCR analysis was performed using the Thermo-Script™ RT-PCR System (Life Technologies, Grand Island, NY, USA) following the manufacturer's instructions. cDNA synthesis was performed using 2 µg of total RNA in the presence of ThermoScript™ reverse transcriptase and oligo( dT) primers at 55℃ for 60 min. Specific primers were designed and synthesized to measure the mRNA expression of the three mouse survivin splice variants and apoptosis genes [12,15]. The primer sets used for the PCR amplification of the target genes were as follows: Survivin 140, 121 and 40 (sense: 5'-TCGCCACCTTCAAGAACTGGCCCTTCCTGGA-3', antisense [splice variants 140 and 40]: 5'-GTTTCAAGAATTCACTGACGGTTAGTTCTT-3' and antisense [splice variant 121]: 5'-GGCTTCTGACAATGCTTG-3'); Bax (For: 5'-TGAGCGAGTGTCT CCGGCGA-3', Rev: 5'-CACGCGGCCCCAGTTGAAGT-3'); Bcl-2 (For: 5'ACCGTCGTGACTTCGCA GAG-3' and Rev: 5'-GTGTGCAGATGCCGGTTCA-3'); p53 (For: 5'-AGAGACCGCCGTACAGAAGA-3' and Rev: 5'-GCATGGGCATCCTTTAACTC-3') and the housekeeping gene β2-micro-globulin (β2m) (sense: 5'-TGACCGGCCTGTATGCTATC-3' and antisense: 5'-CACATGTCTCGATCCCAGTAG-3'). The PCR mixtures were incubated in a DNA thermal cycler (Perkin-Elmer Cetus Instruments, Waltham, MA, USA). The PCR cycle conditions were 29 cycles consisting of denaturation for 30 sec at 94℃, annealing for 30 sec at 58℃ and extension for 30 sec at 72℃. The amplified DNA fragments were separated through electrophoresis on a 1% agarose gel and visualized with ethidium bromide staining. The densities of the ethidium bromide-stained PCR bands were quantified using the Bio-Rad Molecular Imager FX, and the mRNA expression of the investigated genes was normalized against the mRNA expression of β2m.
TUNEL assay
The level of DNA fragmentation, a hallmark of apoptosis, was detected via the terminal transferase-mediated dUTP-biotin nick end-labeling (TUNEL) assay. The reactions were performed using an in situ cell death detection POD kit (Roche Diagnostics, Mannheim, Germany) according to the manufacturer's instructions. Freshly excised testicular tissue was fixed in 10% formalin, embedded in paraffin and sectioned at a thickness of 4 µm. The sections were deparaffinized and rehydrated with serial changes of xylene and ethanol followed by Proteinase K treatment (20 mg/l) (Sigma-Aldrich, St. Louis, MO, USA) for 15 min. The sections were then treated with reaction mixture containing TdT and biotinylated 16-dUTP for 1 h at 37℃. The labeled DNA was visualized with peroxidase-conjugated antidigoxigenin antibody using 3,3'-diaminobenzidine as the chromogen. Randomly selected microscopic fields were used for the evaluation of DNA fragmentation. One hundred seminiferous tubules from each testis were evaluated in circular cross-sections from six testes in each group. The number of TUNEL-positive nuclei per tubule was counted and expressed as the mean±SEM for each group.
Histological and morphometric analyses
Testicular tissue was fixed in Bouin's solution (7.5 ml of saturated picric acid, 2.65 ml of glacial acetic acid and 2.5 ml of 7% formaldehyde) for 24 h, washed in PBS buffer and embedded in paraffin blocks in preparation for microscopic analysis. Four-micron sections from each ipsilateral and contralateral testis were stained with hematoxylin and eosin (H&E) and analyzed under light microscopy. The testicular tissue was evaluated in a random order, and the examiner was blinded to the identity of the animal group. Slides prepared from different portions of each testis were evaluated. The following seminiferous tubule parameters were assessed: the STD, GECT and TBS. Tubular parameters were analyzed as described previously, with some modifications [16,17]. Briefly, approximately 10 sections of seminiferous tubules from ipsilateral and contralateral testes that were round or nearly round were selected randomly from each group and photographed using an Olympus Bx51 light microscope (Olympus PM-C35DX camera, Japan). Using Image-Pro Plus software (Version 6.0.0.260; Serial Number-41 M60032-00032), the average diameter of at least five seminiferous tubules (D) was calculated from each photograph by measuring the maximum height and width of each tubule at two perpendicular axes at a final magnification of 400×. We also measured the major and minor axes of the tubule lumen (d) and calculated the GECT (D-d) [18]. The mean TBS was evaluated with Johnsen's score, in which epithelial maturation in the seminiferous tubules was assigned a score of 0~10 [19].
Statistical analysis
All data were expressed as the mean±SEM. For histological data, a t-test (independent) was performed for each group using SPSS software, and a one-way analysis of variance (ANOVA) was applied to assess significant differences between the three animal groups. The Bonferroni and LSD post hoc tests were performed to determine the individual probability values for multiple comparisons using SPSS software (v17.1). The graphs were drawn using Microsoft Excel software. The gene expression data were analyzed using GraphPad Prism (v5.0). The differences between groups were compared with the Kruskal-Wallis test. p values ≤ 0.05 were considered to indicate statistical significance.
RESULTS
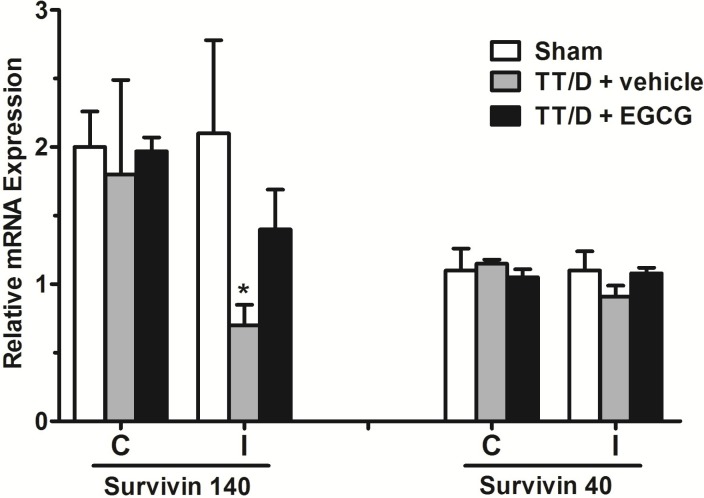
Gene expression of the survivin splice variants
Unlike survivin 121, the mRNA expression of the survivin variants 140 and 40 was detected in the testicular tissue of all three experimental groups (Fig. 1). In the ipsilateral testes, survivin variant 140 expression was significantly down-regulated in the TT/D+vehicle group compared with the sham (0.7±0.15 vs. 2.1±0.68, p<0.05) but was increased upon EGCG treatment (1.4±0.29, p<0.05). In contrast, there was no significant difference in the mRNA expression of survivin 40 between the ipsilateral testes of the sham (1.1±0.14), TT/D+vehicle (0.91±0.08) and TT/D+EGCG (1.08±0.04) mice. The mRNA expression of both survivin variants showed baseline expression in the contralateral testes of the three animal groups.
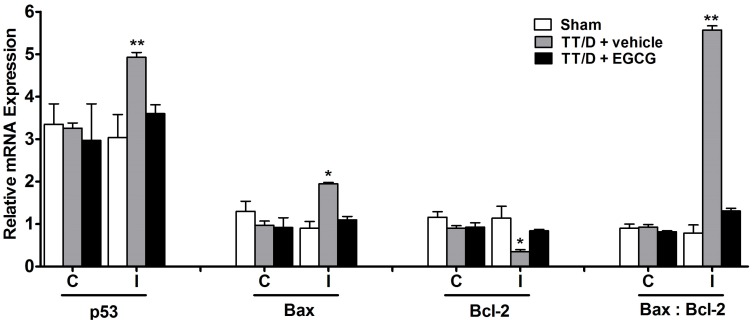
Expression of apoptosis genes
The relative mRNA expression levels of p53, Bax and Bcl-2 are shown in Fig. 2. The expression of the pro-apoptosis gene p53 was significantly elevated in the TT/D+vehicle group compared with the sham levels (4.93±0.11 vs. 3.04±0.54, p<0.01). EGCG administration caused a significant decrease in the p53 mRNA levels of the ipsilateral testes compared with the TT/D+vehicle group (3.6±0.21, p<0.05). Similarly, the mRNA levels of Bax (pro-apoptosis gene) were up-regulated in the ipsilateral testes of the TT/D+vehicle animals compared with the sham (1.95±0.03 vs. 0.9±0.16, p<0.05), which were also significantly suppressed by EGCG treatment (1.1±0.08, p<0.05). Animals treated with EGCG had Bcl-2 (anti-apoptosis gene) expression comparable with sham expression (0.84±0.03 and 1.14±0.28, respectively), which was in contrast to the significantly low Bcl-2 expression in the TT/D+vehicle group (0.35±0.05, p<0.05). The Bax to Bcl-2 ratio was accordingly calculated in the three animal groups, resulting in ratios of 0.79 (0.9/1.14), 5.57 (1.95/0.35) and 1.31 (1.1/0.84) for the sham, TT/D+vehicle and EGCG-treated groups, respectively. The elevated ratio indicates apoptosis induction in the TT/D+vehicle group in comparison to sham and EGCG-treated animals (p<0.01).
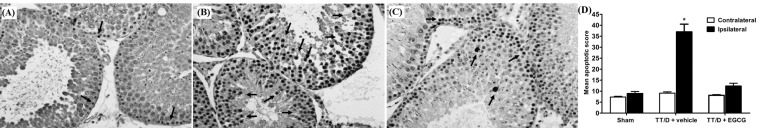
DNA damage in testicular tissue
In the TT/D+vehicle animal group (Fig. 3B), an increased level of DNA damage was observed compared with the sham levels (Fig. 3A) (37±5.1 vs. 8.9±0.94, p≤0.05), as obtained by the TUNEL assay. The TT/D+EGCG animals were protected by EGCG treatment (Fig. 3C) and showed significantly reduced DNA damage compared with TT/D+vehicle animals (12.3±1.31 vs. 37±5.1, p≤0.05).
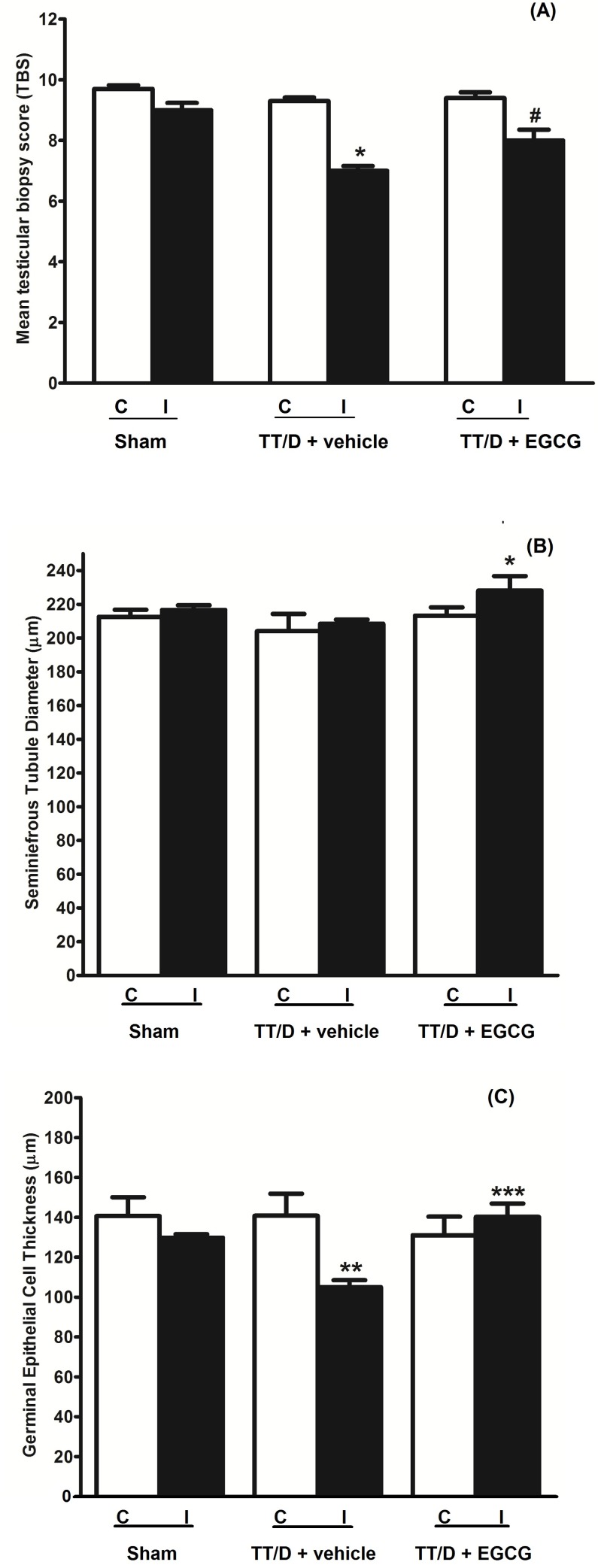
Seminiferous tubule morphometric analysis
Testicular tissue from all three animal groups was stained with H&E to perform the morphometric analysis (Fig. 4). The Johnsen scoring system was used to calculate the TBS for all animal groups (Fig. 4A). The testes from the TT/D+vehicle animals showed significantly lower scores compared with the sham (6.875±0.375 vs. 9.375±0.125, p≤0.0001), indicating damage to spermatogenesis. EGCG treatment exerted a protective effect on spermatogenesis, with a score of 8.313±0.120, which was significantly higher than the TT/D+vehicle score (p≤0.05). The mean TBS for the contralateral testes of the sham, TT/D+vehicle and EGCG-treated groups was 9.7±0.12, 9.3±0.12, and 9.4±0.19, respectively.
The STD of the contralateral testes from the sham (212.64±4.19 µm), TT/D+vehicle (204.08±10.26 µm) and EGCG-treated (213.34±4.86 µm) testes showed no significant difference (Fig. 4B). Interestingly, no significant difference was observed between the sham and TT/D+vehicle mice in the ipsilateral testes (216.72±2.75 µm vs. 208.41±2.5 µm, p>0.05). However, a marked increase in the STD of EGCG-treated ipsilateral testes was found (228.16±8.55 µm) compared with the sham and TT/D+vehicle groups (p<0.02).
The calculated GECT in the contralateral testes of the sham, TT/D+vehicle and TT/D+EGCG animals was 140.68±9.42 µm, 140.8 2±11.05 µm, and 131.06±9.32 µm, respectively (Fig. 4C). A significant decrease in the GECT of the ipsilateral testes was observed in the TT/D+vehicle group compared with the sham group (105.01±3.53 µm vs. 129.82±1.70 µm, p<0.005). In contrast, EGCG treatment significantly increased the mean GECT in the ipsilateral seminiferous tubules compared with the TT/D+vehicle tubules (140.24±6.70 µm vs. 105.01±3.53 µm, p<0.0001). There was no significant difference in the mean GECT of the contralateral testes in the different groups.
DISCUSSION
Survivin is prominently expressed in highly proliferative areas within normal tissues, such as the thymus, testis and hematopoietic cells, with a key regulatory role in apoptosis and mitosis [20-23]. Thus, the inactivation of survivin expression in normal cells appears to have detrimental effects. However, the regulation of survivin expression and, consequently, its function presents some complexity. Transcription of the survivin gene results in three alternatively spliced survivin mRNAs in humans, mice and chickens, each with different expression patterns and different abilities to prevent apoptosis [24]. The molecular mechanisms through which survivin, particularly the survivin splice variants, control cell division and inhibit apoptosis have been extensively explored but are not yet fully understood [25,26]. As a result of their key role in different human cancers, most of the current studies have focused on the role of survivin and its splice variants as prognostic markers in carcinogenesis [27,28]. However, little attention has been given to their role in non-cancer conditions. One study characterized their role in cryptorchidism-induced apoptosis, revealing decreased expression of murine survivin variants 140 and 40 mRNA accompanied by reduced survivin protein expression [13]. Another study also showed that murine survivin variants 140 and 40 were involved in the regeneration of nerve and/or spinal cord injuries [14]. In both studies, the expression of murine survivin splice variant 121 was undetectable, a finding that is further supported by this study. To shed light on the role of the survivin splice variants during TT/D-induced apoptosis, we evaluated their gene expression in a TT/D murine model. Survivin variant 140, the most predominant survivin splice variant, showed a marked decrease in its gene expression as a result of TT/D. Conversely, survivin variant 40 expression was insignificantly changed in response to TT/D, and there was no detectable expression of survivin variant 121. The decreased survivin 140 expression was restored by EGCG treatment, which was also associated with normal morphological features of the seminiferous tubules and spermatogenesis.
Testicular tissue homeostasis is maintained through a fine balance between cellular proliferation and germ cell apoptosis (GCA) [29]. Achieving this balance, especially in the testis, ensures the integrity of the genetic material and its dissemination, in which survivin plays a central role. Survivin phosphorylation by p34cdc2-cyclin B1 has been associated with the activation of the spindle assembly check point, which couples survivin with apoptosis [30-32]. Survivin has also been reported to be highly expressed during the long first prophase of spermatocytes, suggesting a major role in meiosis [21,33]. However, whether survivin simultaneously acts as an IAP in the maintenance of homeostasis is still unclear. Low cytoplasmic survivin levels were detected in meiotically dividing germ cells, which are susceptible to apoptosis, especially during metaphase [34-36]. This finding could be due to the chromosomal localization of survivin during mitosis. In our study, low survivin 140 levels were observed in untreated TT/D mice, which coincided with the lowered TBS. A low TBS indicates a lack of spermatozoa and late spermatids with the presence of many early spermatids, which could suggest that survivin might be acting as an IAP and that its low levels induce GCA under this condition. Decreased survivin levels during TT/D could also be proposed to lead to the activation of the spindle assembly check point, thereby inducing apoptosis.
The relationship between EGCG and survivin expression has been investigated in several studies. EGCG is recognized as a chemopreventive agent, which is inversely correlated with the prevalence of various cancer types, including breast, prostatic, colonic, mammary, ovarian and pancreatic cancers [37-39]. In most of these cancers, survivin has been shown to be a molecular target of EGCG, in which increased survivin levels are suppressed by EGCG treatment [40-42]. Nonetheless, in non-cancer diseases, the relationship between EGCG and the expression of survivin splice variants has not been reported prior to this study. Our results indicate that in non-cancer pathological conditions, EGCG up-regulates survivin expression toward reestablishing it at normal levels. Therefore, EGCG may act through different mechanisms to modulate the expression of survivin in both cancer and non-cancer conditions.
Based on earlier studies, we suggest that signaling pathways of oxidative stress, inflammation and apoptosis are induced in the setting of TT/D [43]. A common regulator in these pathways is NFκB, which is up-regulated in such pathways in non-cancer conditions [44]. NFκB activation has been associated with the up-regulation of p53 and Bax expression but the down-regulation of Bcl-2 and survivin expression [45,46]. In addition, an IAP-IAP complex between survivin and XIAP was shown to increase the inhibition of caspase 3 activity and degradation of NFκBIa, with the consequent activation of NFκB [47,48]. EGCG has been reported to exert its anti-apoptotic activity via the inactivation of NFκB, caspase 1 and caspase 3 [49]. Thus, during TT/D, EGCG can be hypothesized to promote GCA survival through the inactivation of the NFκB pathway, which is associated with the down-regulation of Bcl-2, increased survivin expression and its concomitant action with XIAP. In our study, we also investigated the role of other apoptotic markers, both anti- and pro-survival, during TT/D-induced apoptosis. Hypothetically, such markers can be used to evaluate the level of infertility and fertilizability of spermatozoa and also serve as diagnostic factors [50]. For example, at the molecular level, p53 gene expression and DNA fragmentation can be considered good markers for sperm survival and the ability to fertilize, whereas an increased ratio of Bax to Bcl-2 gene expression can serve as an important marker for apoptosis induction [50]. These findings are supported by our previous and current findings, in which the Bax to Bcl-2 ratio and p53 expression were increased in parallel with increased damage to spermatogenesis and DNA fragmentation. The latter has been associated with low sperm motility [51], impaired fertility, disordered embryonic development, high rates of miscarriage and an increased risk of defects in the offspring [52].
The effect of EGCG on the structure of seminiferous tubules was also noticeable. EGCG treatment was able to protect both the TBS and GECT from TT/D injury and prevent damage to spermatogenesis. An interesting finding was that although TT/D injury did not affect the STD, EGCG treatment led to a significant increase in STD. This result might indicate that EGCG treatment associated with the restoration of normal survivin 140 expression could have contributed to the enhancement of germ cell survival and proliferation. This result could also suggest that survivin might be acting as a pro-survival molecule and mitotic checkpoint regulator in the testis in the presence of EGCG.
In conclusion, this study showed, for the first time, that the protective role of EGCG was associated with the expression of survivin 140 but not variants 121 and 40. However, establishing the role of survivin 140 as an IAP or a cell cycle regulator in this condition requires further investigation. Additionally, EGCG was found to enhance the homeostasis of testicular tissue and spermatogenesis and protect it from TT/D-induced damage.
 XML Download
XML Download