

 PDF
PDF ePub
ePub Citation
Citation Print
Print


ABBREVIATIONS
ANS
1-anilinonaphthalene-8-sulfonic acid
BSA
bovine serum albumin
PBS
phosphate-buffered saline
Py-3-Py
1,3-di(1-pyrenyl)propane
RET
radiationless energy transfer
SPMV
synaptosomal plasma membrane vesicles isolated from bovine cerebral cortex
SPMVTL
liposome of total lipids extracted from SPMV
SPMVPL
liposome of phospholipids extracted from SPMV
INTRODUCTION
Two general theories have been proposed to explain the action of local anesthetics on the sodium channel. The first considers the direct binding of local anesthetic molecules to the specific receptors on the sodium channels [1-4], and the second proposes the general perturbation of the bulk membrane structure by anesthetics and its consequences on the channel function [3-9]. A significant amount of evidence supports the specific receptor hypothesis [10]. General membrane perturbation can also explain the anesthetic actions [10]. On the other hand, the precise location of the molecular mechanism of action of local anesthetics is still a subject of controversy.
The ability to regulate the cell volume is one of the elementary requirements of cell survival, and volume regulatory mechanisms are a integral part of the intracellular cascade responsible for transmitting hormonal signals [11,12]. The cell volume is linked to the ion channels and carriers at the cell membrane, and to the cellular metabolic activity [11,12]. The cell volume is an integral element within the cellular machinery and regulates the cellular performance. The role of the cell volume in cell functions involves the erythrocyte volume and shape, epithelial transport, the regulation of the metabolism, receptor recycling, hormone and transmitter release, excitability and contraction, migration, pathogen host interactions, cell proliferation, cell death etc. [12].
The effects of local anesthetics on motion, order and phase transitions of bulk bilayer systems of native or model membranes have attracted considerable attention over the past few decades. This is due to the interest in biological membranes as well as the unique information on the intermolecular interactions that can be derived from an investigation of the volume changes. The potency of an anesthetic increases roughly in proportion to its lipid/water partition coefficient, strongly suggesting an amphiphilic site for anesthetic molecules [3,4,8,13-15]. Yun et al. [8] reported that local anesthetics decreased the microviscosity of synaptosomal plasma membrane vesicles isolated from the bovine cerebral cortex (SPMV). In addition, differential scanning thermograms of dimyristoylphosphatidylcholine multilamellar liposomes show that local anesthetics decreased the phase transition temperature significantly, broadened the thermogram peaks, and reduced the size of the cooperative unit.
Little attention has been paid to the effects of lidocaine·HCl on the thickness of biological and model membranes. The aim of this study was two-fold: (i) to provide a basis for studying the mechanism of action of local anesthetics, and (ii) to develop a fluorescence quenching method to determine the membrane thickness. In the present study, the effect of lidocaine·HCl on the thickness of neuronal and model membrane lipid bilayers was examined using the energy transfer between the surface fluorescent probe, 1-anilinonaphthalene-8-sulfonic acid (ANS), and the hydrophobic fluorescent probe, 1,3-di(1-pyrenyl) propane (Py-3-Py).
METHODS
Materials
The fluorescent probes, Py-3-Py and ANS, were purchased from Molecular Probes, Inc. (Junction City, OR, USA). Lidocaine·HCl, N-2-hydroxyethyl-piperazine-N'-2-ethanesulfonic acid (Hepes) and bovine serum albumin (BSA) were supplied by Sigma Chemical (St. Louis, MO, USA). All other reagents obtained commercially were of the highest quality available. Water was deionized.
Liposomes preparation
The total lipids were extracted from the SPMV, as described previously [14]. The cholesterol content of the extracted total lipids was determined using the Liebermann-Buchard reaction [22]. The phospholipids were quantified by measuring the amounts of inorganic phosphate [23] after hydrolysis of the phospholipids at 180℃ in 70% HClO4 [24]. The SPMV had a high lipid to protein ratio (0.942 mg total lipids/1 mg protein) and a low cholesterol to phospholipid molar ratio (0.593±0.011: cholesterol 0.208±0.010, phospholipids 0.702±0.025). For the calculation, an average molecular weight of 775 for phospholipids is assumed and the molecular weight of cholesterol is 387. The phospholipids were composed (mol%) of phosphatidylcholine (41.55±0.91), phosphatidylethanolamine (36.83±0.48), phosphatidylserine (13.60±0.26), sphingomyelin (4.15±0.16), phosphatidylinositol (2.90±0.09) and lysophosphatidylcholine (0.97±0.03).
Stock solutions of total lipids and phospholipids were prepared in chloroform. The concentration of the phospholipid stock solutions was 0.2 mg/ml. Giant unilamellar vesicles (GUVs: SPMVTL and SPMVPL) with a mean diameter of 45 µm were prepared using the method reported by Angelova and Dimitrov [25,26] and Angelova et al. [27]. A special temperature-controlled chamber described previously [28,29] was used to grow the GUVs. The experiments were carried out in the same chamber after vesicle formation, using an inverted microscope (Axiovert35: Zeiss, Thornwood, NY). The following steps were used to prepare the GUVs. Approximately 3 µl of the lipid and phospholipid stock solution was spread on each Pt wire under a N2 stream. To remove the residues of the organic solvent, the chamber was placed in a liophilizer for ~2 h. To add the aqueous solvent inside the chamber (Millipore water 17.5 MΩ/cm), the bottom part of the chamber was sealed with a coverslip. Millipore water was previously heated to the desired temperature (80℃), and sufficient water was then added to cover the Pt wires. Immediately after this step, the Pt wires were connected to a function generator (Hewlett-Packard, Santa Clara, CA), and a low-frequency AC field (sinusoidal wave function with a frequency of 10 Hz and an amplitude of 3 V) was applied for 90 min. After vesicle formation, the AC field was turned off.
Data analysis
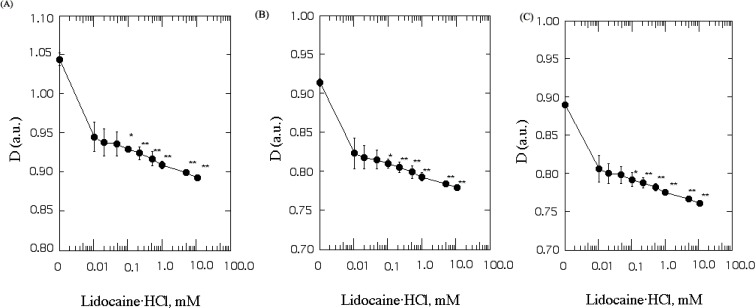
Each value is expressed as the mean±SEM (n=5 for all experiments). The differences in D and the a.u. (thickness) values were analyzed statistically using a Student's t-test. The asterisk and double asterisks denote p<0.05 and p<0.01, respectively.
Determination of thickness of SPMV, SPMVTL and SPMVPL
All fluorescence measurements were obtained with a Multi Frequency Cross-Correlation Phase and Modulation Fluorometer (Model; ISS K2-003). The cuvette temperature was maintained at 37.0±0.1℃ in a circulating water bath (pH 7.4). The bandpass slits were 10 nm on excitation and 5 nm on emission. Blanks, which were prepared under identical conditions without fluorescent probes, were used as the controls.
The effect of lidocaine·HCl on the thickness of neuronal and model membrane lipid bilayers was determined using energy transfer between the surface fluorescent probe ANS and the hydrophobic fluorescent probe, Py-3-Py. Py-3-Py was excited at 330 nm, and the monomer fluorescence intensity of Py-3-Py was read at 379 nm. Subsequently, 30 µM ANS (60 µl of 10-3 M solution in water) was added. After 1 min, the Py-3-Py monomer fluorescence was measured again. ANS was located on the membrane surface. This concentration was chosen because it produced 50% quenching of the Py-3-Py monomer fluorescence in SPMV, SPMVTL and SPMVPL. The efficiency of Py-3-Py quenching by ANS was calculated using equation 1, considering the Py-3-Py monomer and ANS as the donor and acceptor, respectively.
where Id and Ida represent the fluorescence intensity of the donor (Py-3-Py monomer) in the absence and presence of acceptor (ANS), respectively. The membrane thickness was considered to be proportional to the distance (D) between the donor and acceptor, and can be calculated as follows:
where Φ is the number of acceptor/volume and <E> is the average efficiency of radiationless energy transfer (RET) [30]. The following modified equation was used to calculate D:
where F' is the fluorescence intensity of the concentration of acceptor used in the RET experiments, and Fmax is the maximal fluorescence of the acceptor in the membrane (both values were obtained by ANS binding studies in the absence of Py-3-Py). The ratio, F'/Fmax, provides a measure of the binding of ANS to the membrane surface and provides a correction for the effects of lidocaine·HCl on this binding.
ANS (2 µl of 10-2 M solution) was added repeatedly to the sample at 15 sec intervals and the measurements were made as soon as the probe was added. In each case, at least 14 concentrations of the probe were used. The dissociation constants and Fmax values for ANS-membrane complex were calculated from the double-reciprocal plots by linear regression analysis. The correlation coefficients were >0.99.
RESULTS
Molecular movement eventually increases the molecular volume. On the other hand, it is not known how much molecular movement causes an increase in molecular volume. This study measured how much of the membrane thickness (D) was decreased (or increased) by lidocaine·HCl taking advantage of the fluorescence quenching technique.
In the present conclusion, it would be rational to use liposome comprised of the total lipids extracted from neuronal membranes and liposome, which is made up of phospholipids extracted from neuronal and model membranes, as samples in a study of the structure-activity relationship of local anesthetics.
The purity of SPMV
The purity of SPMV was assessed using the enzymatic and morphological criteria. The specific activities of Na+, K+-ATPase, acetylcholinesterase and 5'-nucleotidase were enriched approximately 4-, 2.5- and 3-fold, respectively, in the plasma membrane fraction with respect to the crude homogenates. Transmission electron microscopy of the SPMV indicated very high purity [16]. The vesicles, which were separated according to size, showed a homogeneous distribution and no intracellular organelles or leakage.
Effect of lidocaine·HCl on the thickness (D) of SPMV, SPMVTL and SPMVPL
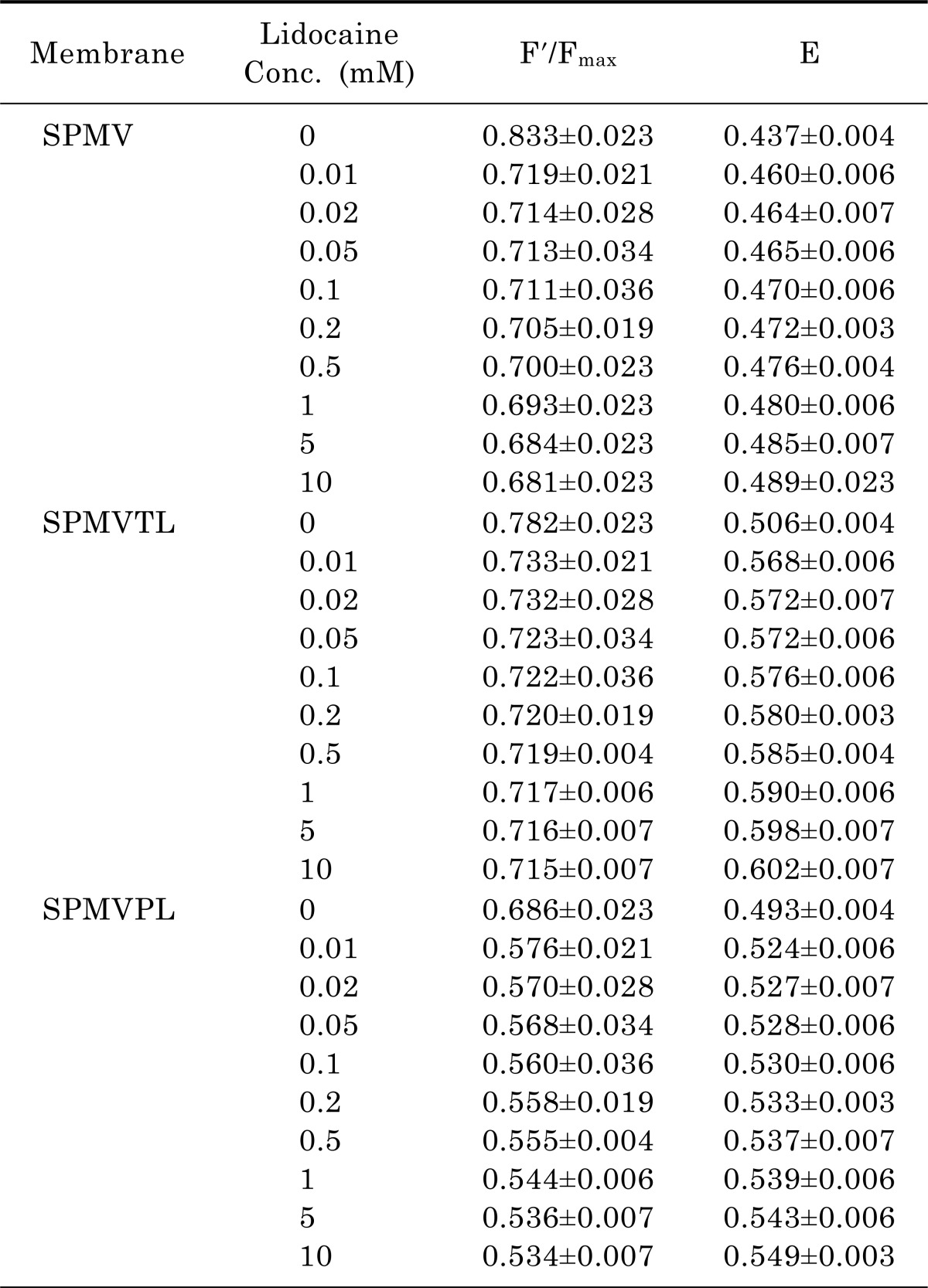
The membrane thickness (D) was measured by energy transfer from Py-3-Py to ANS in SPMV, SPMVTL and SPMVPL. The intact neuronal and model membrane thickness (D) in arbitrary units was 1.044±0.008, 0.914±0.005 and 0.890±0.003 (37℃, pH 7.4), respectively (Table 1). Table 2 lists the F'/Fmax and E values. Lidocaine·HCl decreased the thickness (D) in a dose-dependent manner (Fig. 1) (37℃, pH 7.4). Moreover, significant decreases in the thickness (D) by lidocaine·HCl was observed, even at 0.1 mM and above, respectively (Fig. 1). This suggests that lidocaine·HCl decreases the thickness (D) of the SPMV, SPMVTL and SPMVPL lipid bilayer, which means that the membrane has been expanded.
DISCUSSION
The volume of matter changes in proportion to the molecular movement. This is self-evident from the fact that when water becomes vapor, its volume increases. From previous results, lidocaine·HCl increased the range and rate of lateral mobility of the bulk (inner+outer monolayer) SPMV, SPMVTL and SPMVPL lipid bilayers at concentrations as low as 0.02 mM (n=5, p<0.05) [31]. In addition, lidocaine·HCl increased the rotational mobility of bulk SPMV, SPMVTL and SPMVPL lipid bilayers at 0.1, 0.2 and 0.1 mM, respectively [31]. Furthermore, lidocaine·HCl increased the annular lipid fluidity in the SPMV lipid bilayers, and the local anesthetics induced protein clustering in SPMV [31].
Park et al. [31] clearly shows that lidocaine·HCl increases the bulk lateral and rotational mobility of the neuronal and model membrane lipid bilayers. The decreased thickness of SPMV, SPMVTL and SPMVPL lipid bilayers by lidocaine·HCl was attributed to the lateral and rotational mobility of the SPMV, SPMVTL and SPMVPL lipid bilayers, and the annular lipid fluidity was increased by lidocaine·HCl. In addition, the fact that the thickness of the SPMV, SPMVTL and SPMVPL lipid bilayer decreases in a concentration-dependent manner strongly suggests that the lateral and rotational mobility of the SPMV, SPMVTL and SPMVPL lipid bilayers was increased by lidocaine·HCl. This not only signifies an expansion of the SPMV, SPMVTL and SPMVPL lipid bilayer but also suggests an interdigitation phenomenon between the inner and outer monolayers. Therefore, it is presumed that lidocaine·HCl causes interdigitation in the lipid bilayers of SPMV, SPMVTL and SPMVPL. Therefore, the effects of lidocaine·HCl on the neuronal and model membranes are due to its direct and indirect actions.
The membrane thickness (D) of the neuronal and model membrane lipid bilayers was not changed by 0.02 mM lidocaine·HCl. When it reaches a concentration of 0.1 mM lidocaine·HCl, the thickness (D) of the membrane lipid bilayers shows a significant decrease. This means that the thickness (D) of the lipid bilayers is affected only by a considerable change in the lipid mobility of neuronal and model membranes. If not, the fluorescence probe technique used in this study would not detect the infinitesimal changes in membrane thickness (D).
Opinions have been divided as to whether local anesthetics interfere with the membrane protein function by binding directly to the proteins, or whether the main modes of action occur indirectly through a change in the physicochemical properties of the lipid membranes into which the local anesthetics readily diffuse. Because biological membranes have a complex composition, it was not feasible to simultaneously monitor the changes in the local lipid environment and determine its effect on the membrane protein function. The multiplication effects can be explained by citing the increased mobility of protein triggered by lipids, but the reverse case of protein triggering a change in lipids is more likely. Nevertheless, local anesthetics increase the mobility of the neuronal lipid bilayers but the direct effects of local anesthetics on the protein appear to have magnified these effects on the lipid. Specifically, before, during or even after the interaction of the local anesthetics with the sodium channels, the fluidization of membrane lipids might provide an ideal microenvironment for optimum local anesthetic effects. In conclusion, these results suggest that local anesthetics, in addition to their direct interaction with sodium channels, interact concurrently with the membrane lipids, affecting the fluidity of the neuronal membrane lipid bilayers.

 XML Download
XML Download