

 PDF
PDF ePub
ePub Citation
Citation Print
Print


ABBREVIATIONS
AP-1
activator protein-1
COX-2
cyclooxygenase-2
ERK
extracellular signal-related kinase
GAPDH
glyceraldehyde 3-phosphate dehydrogenase
IL-1β
interleukin-1β
IL-6
interleukin-6
iNOS
nitric oxide synthase
JNK
C-jun N-terminal kinase
LPS
lipopolysacharide
TLR-4
Toll-like receptor-4
MAPK
mitogenactivated protein kinase
NF-κB
nuclear factor-kappa B
NO
nitric oxide
PARP
poly ADP-ribose polymerase
INTRODUCTION
Developing anti-inflammatory agents from natural or synthetic products is prudent because the inflammatory response is involved in various conditions including cancer, obesity, aging, atherosclerosis and neurodegeneration [1,2]. An uncontrolled and prolonged inflammatory reaction may underlie the pathogenesis of chronic diseases [1]. We recently reported the prowess of ailanthoidol, a benzofuran-type neolignan containing alcoholic and phenolic hydroxyl groups, in curtailing various inflammatory reactions in vitro and in vivo [3]. Ailanthoidol also exhibits several pharmacological effects that include inhibition of tumor promotion induced by 12-O-tetradecanoyl-phorbol-13-acetate [4], and prohibiting copper ion-mediated oxidation of low-density lipoprotein [5] and antimicrobial activity [6]. Various organic synthesis methods of ailanthoidol have been reported [3,7,8].
Inflammation is a protective effort by an organism to eradicate injurious stimuli and initiate healing. An inflammatory reaction directs immune system components to the site of injury or infection, and is manifest by increased blood supply and vascular permeability, which allows chemotactic peptides, neutrophils and mononuclear cells to leave the intravascular compartment [9]. Various infecting agents, such as bacteria and pro-inflammatory cytokines, can activate macrophages, which are critical immune cells to regulate innate immunity, through certain receptors [10]. The interaction of Toll-like receptor (TLR)-4 with the ligand, lipopolysaccharide (LPS), induces an intracellular signaling cascade that activates the mitogen-activated protein kinase (MAPK) family, extracellular signal-related kinase (ERK), p38, C-jun N-terminal kinase (JNK) and adaptor molecules, and ends in the activation of key pro-inflammatory transcription factors, such as activator protein (AP)-1 and nuclear factor-kappa B (NF-κB) [10,11]. Eventually, these signaling events lead to the transcriptional activation of macrophages and subsequent expression of pro-inflammatory genes including inducible nitric oxide synthase (iNOS), cyclooxygenase (COX)-2 and various cytokines [10-12].
During our on-going screening program designed to identify the anti-inflammatory potential of natural and synthetic compounds, we have prepared several ailanthoidol derivatives (compounds 1~6). Our previous report demonstrated that ailanthoidol has anti-inflammatory activities due to the inhibition of iNOS and COX-2 expression and impairment of the release of various inflammatory cytokines [3]. To evaluate whether ailanthoidol derivatives regulate the inflammatory response, we presently investigated the anti-inflammatory effects of these derivatives using LPS-induced inflammatory responses.
METHODS
Chemicals and reagents
Ailanthoidol and its derivatives (Fig. 1) were synthesized using Sonogashira coupling, iodine induced cyclization and Wittig reaction as previously reported [13]. LPS derived from Escherichia coli, dimethylsulfoxide (DMSO) and anti-β-actin antibody were obtained from Sigma-Aldrich (St Louis, MO, USA). Dulbecco's modified Eagle's medium (DMEM), fetal bovine serum (FBS), penicillin and streptomycin were obtained from Hyclone (Logan, UT, USA). Cell Titer 96 Aqueous One Solution and Griess reagent system were purchased from Promega (Madison, MI, USA). Prostaglandin (PG) E2 enzyme-linked immunosorbent assay (ELISA) kits were obtained from ENZO Life Sciences (Farmingdale, NY, USA). Interleukin (IL)-1β and IL-6 ELISA kits were purchased from eBioscience (San Diego, CA, USA). Anti-iNOS, anti-COX-2, anti-inhibitor of NF-κB (IκB-α), anti-phospho-ERK1/2, anti-ERK1/2, anti-phospho-JNK, anti-JNK, anti-phospho-p38, anti-p38, anti-phospho-c-Jun, anti-c-Jun and anti-poly ADP-ribose polymerase (PARP) antibody were obtained from Cell Signaling Technology (Danvers, MA, USA).
Cell culture and cell viability assay
RAW264.7 murine macrophages obtained from the Korean Cell Bank (Seoul, Korea) were cultured in DMEM containing 10% FBS, 100 U/ml penicillin and 100 µg/ml streptomycin at 37℃ in 5% CO2. The effects of various ailanthoidol derivatives on cell viability were tested using the CellTiter 96® AQueous One Solution Assay of cell proliferation. RAW264.7 cells were plated at a density of 2×104 cells in a 96-well flat-bottom plate and ailanthoidol derivatives were added to each plate at the indicated concentrations. After a 24 h incubation period, the number of viable cells was counted according to the manufacturer's instructions. The optical densities at 490 nm were measured using a microplate reader (Tecan, San Jose, CA, USA).
Measurement of nitrite, PGE2 and cytokines
The amount of nitrite, PGE2, IL-1β and IL-6 produced by mouse macrophages was measured in RAW264.7 cell culture supernatant. RAW264.7 cells were plated at a density of 2.5×105 cells in a 48-well cell culture plate with 500 µl of culture medium and incubated for 12 h. They were then treated with indicated concentrations of ailanthoidol derivatives plus LPS (100 ng/ml) and incubated for another 24 h. The amount of nitrite was measured using the Griess reagent system according to the manufacturer's instructions. PGE2, IL-1β and IL-6 were measured using ELISA according to the manufacturer's instructions.
Quantitative real-time reverse-transcription polymerase chain reaction (qRT-PCR)
Total RNA was isolated from RAW264.7 cells using Trizol Reagent (Invitrogen, Carlsbad, CA, USA). DNA was eliminated from total RNA using RNA Qualified RNase-Free DNase (Promega) and cDNA was synthesized by GoScript™ Reverse Transcription System (Promega). qRT-PCR assay was carried out with LightCycler (Roche Diagnostics, Basel, Switzerland) using LightCycler FastStart DNA Master SYBR Green I (Roche Diagnostics). All the experiments were repeated twice and in triplicate each time. Transcripts of glyceraldehyde 3-phosphate dehydrogenase (GAPDH), as a housekeeping gene, were quantified as endogenous RNA of reference to normalize each sample. Relative quantities were estimated by the delta-delta-Ct method. The results were normalized as relative expression in which the average value of the iNOS, COX-2, IL-1β and IL-6 mRNA was divided by the average value of GAPDH mRNA. The primer used in this study were murine iNOS: F 5'-CCT CCT CCA CCC TAC CAA GT-3', R 5'-CAC CCA AAG TGC TTC AGT CA-3'; murine COX-2: F 5'-AAG ACT TGC CAG GCT GAA CT-3', R 5'-CTT CTG CAG TCC AGG TTC AA-3'; murine IL-1β: F 5'-TTC TCC ACA GCC ACA ATG AG-3', R 5'-ACG GAC CCC AAA AGA TGA AG-3; murine IL-6: F 5'-CAT CCA GTT GCC TTC TTG GGA-3', R 5'-CCA GTT TGG TAG CAT CCA TC-3'; GAPDH: F 5'-TCT TGC TCA GTG TCC TTG C-3', R 5'-CTT TGT CAA GCT CAT TTC CTG G-3'.
Western blotting analysis
Whole cell lysates and nuclear protein were extracted using PRO-PREP Protein Extraction Solution (iNtRON Biotechnology, Seongnam-Si, Korea) and a Nuclear extraction kit (Millipore, Billerica, MA, USA) according to the manufacturer's instructions. Whole cell lysates and nuclear protein (30 µg protein/lane) were separated by 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE). The separated proteins were electrophoretically transferred onto nitrocellulose membranes. Immunoreactive bands were detected by incubating the samples with horseradish peroxidase (HRP)-conjugated secondary anti-bodies and visualized using a WEST-ZOL plus Western Blot Detection System (iNtRON Biotechnology) and Davinch-Chemi CAS-400SM Western Imaging System (Davinch-K, Seoul, Korea). The density of bands was assessed by Total Lab software (Davinch-K).
Statistical analysis
The data are depicted as the mean±SEM. The values were evaluated by one-way analysis of variance (ANOVA) with Bonferroni multiple comparison post tests using the GraphPad Prism 4.0 software (GraphPad Software Inc., San Diego, CA). A p-value<0.05 was considered to be statistically significant.
RESULTS
Compound screening
To examine the potential anti-inflammatory activity of the ailanthoidol derivatives, we used murine macrophage RAW264.7 cells, which are an established excellent model for anti-inflammatory drug screening. We first assayed the ability of the ailanthoidol derivatives, compounds 1~6 (Fig. 1), to decrease NO release after LPS stimulation because LPS induces iNOS and then NO release. RAW264.7 cells were incubated with LPS and a range of concentrations (1~50 µM) of ailanthoidol derivatives for 24 h. Then, culture media were harvested and nitrite levels were measured. All derivatives were able to inhibit NO production. Compound 4 was most potent, with an IC50 of 4.38 µM (Table 1).
Compound 4 inhibits NO and PGE2 release
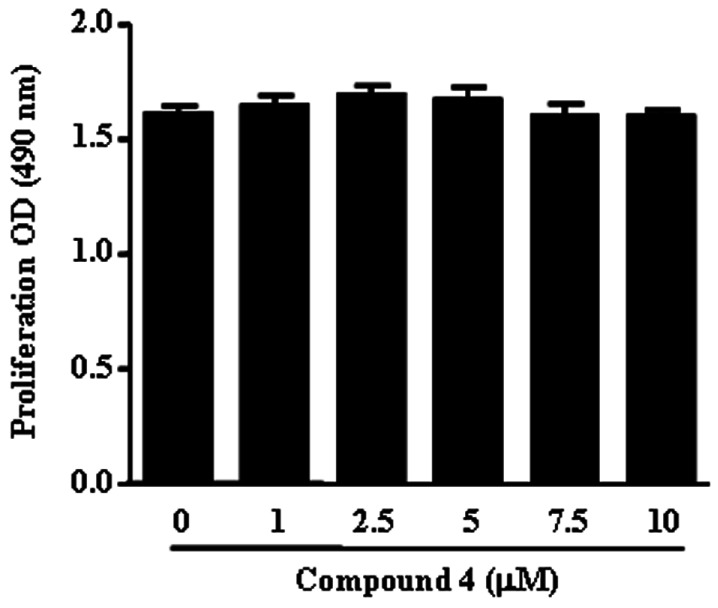
Since compound 4 displayed the lowest IC50 on the inhibition of NO release, this compound was selected for further investigation. To rule out that the inhibitory effect on NO release was due to cytotoxicity, cell viability was analyzed. Compound 4 displayed no toxic effects at concentrations of 10 µM (Fig. 2). We used up to 10 µM compound 4 for the remaining experiments.
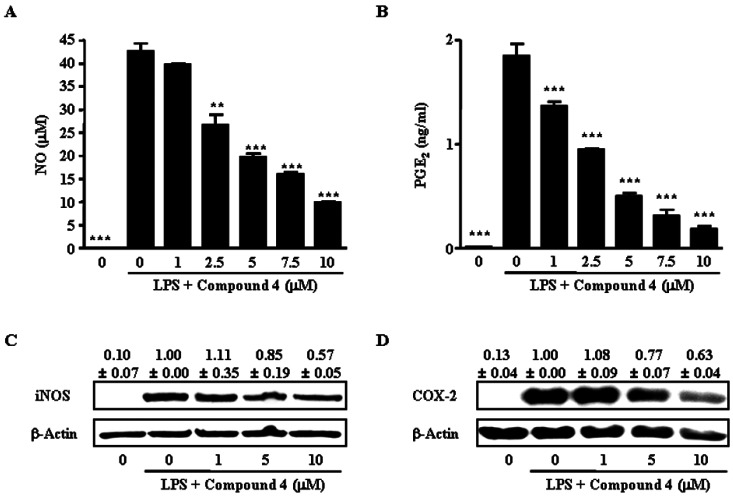
The next experiment determined the effect of compound 4 on the LPS-induced release of the inflammatory mediators, NO and PGE2, by RAW264.7 cells. As shown in Fig. 3A and 3B, compound 4 inhibited LPS-induced NO and PGE2 production in a dose-dependent manner. The nitrite concentration in LPS-stimulated cells and in those exposed to 10 µM compound 4 was 42.5±1.8 µM and 9.9±0.2 µM, respectively. The inhibitory effects of compound 4 on PGE2 production in LPS-exposed cells were similar to their effects on NO production (Fig. 3B). The PGE2 concentration in LPS-stimulated cells and in those exposed to 10 µM compound 4 was 1.85±0.16 ng/ml and 0.19±0.04 ng/ml, respectively.
To determine the mechanism by which compound 4 reduced LPS-induced NO and PGE2 production, the effect of compound 4 on iNOS and COX-2 protein expression in RAW264.7 cells was studied using Western blot analysis, since those enzymes catalyze the reaction for NO and PGE2, respectively. Consistent with the findings related to NO and PGE2 releases, the protein expression of iNOS and COX-2 induced by LPS in RAW264.7 cells were also reduced by compound 4 treatment (Fig. 3C and 3D).
Compound 4 blocks the release of pro-inflammatory cytokines
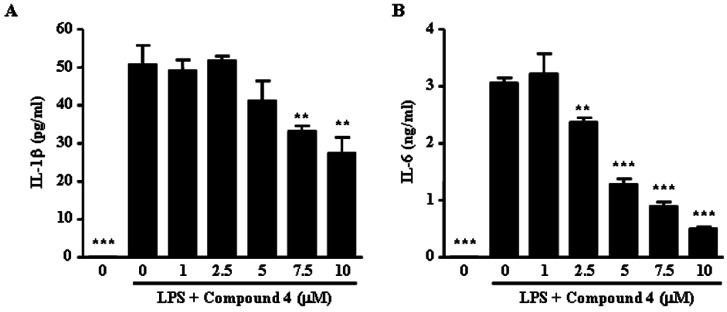
We next examined if compound 4 reduced the release of pro-inflammatory cytokines in LPS-stimulated RAW264.7 cells. Although the concentrations of IL-1β and IL-6 were not detected in vehicle-treated RAW264.7 cells, LPS treatment elevated the levels of IL-1β (55.26±3.90 pg/ml) and IL-6 (3.05±0.14 ng/ml) in LPS-treated RAW264.7 cells. Compound 4 induced marked suppression of the increases induced by LPS in these cytokines (Fig. 4). LPS-treated RAW264.7 cells exposed to compound 4 at concentrations of 1, 5 and 10 µM displayed a dose-dependent inhibited production of IL-1β (13.2±8.6%, 27.3±16.1% and 51.5±12.5%, respectively) and IL-6 production (0%, 58.4±6.0% and 83.8±1.6%, respectively). These results indicate that compound 4 suppressed various inflammatory mediators including NO and PGE2, as well as pro-inflammatory cytokines, such as IL-1β and IL-6.
Compound 4 suppresses the mRNA expression of iNOS, COX-2, IL-1 β and IL-6
Since compound 4 suppressed the protein levels of iNOS, COX-2 and pro-inflammatory cytokines in LPS-stimulated RAW264.7 cells, qRT-PCR was used to assess the effects of compound 4 on LPS-induced gene expression of iNOS, COX-2, IL-1β and IL-6 in RAW264.7 cells. Upon LPS treatment, the mRNA expressions of these four genes were markedly augmented. The concentration response for inhibition of iNOS, COX-2, IL-1β and IL-6 mRNA expressions is shown in Table 2. The effect of compound 4 on mRNA expression of iNOS and COX-2, as well as IL-1β and IL-6, was coincident with protein expression and secretion in supernatants.
Compound 4-reduced inflammatory mediator release is regulated by MAPK and AP-1
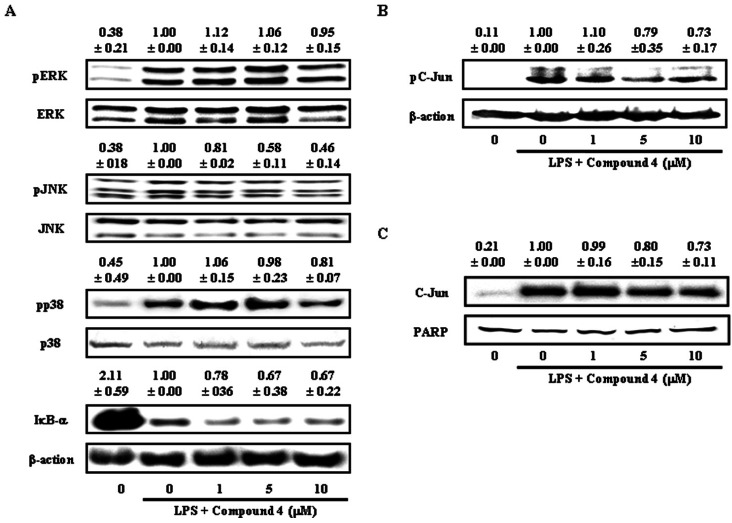
To ascertain the effect of compound 4 on the pathways involved in regulating the expressions of iNOS, COX-2 and pro-inflammatory cytokines, degradation of IκB-α, an important biochemical event for the nuclear translocation of NF-κB, and phosphorylation of MAPK molecules were analyzed by Western blotting. Phosphorylation of JNK was inhibited by compound 4 treatment, where inhibitory action was dose-dependent on JNK phosphorylation (Fig. 5A). However, the effect of compound 4 on the phosphorylation of ERK and p38 was not significant. In addition, as shown in Fig. 4A, compound 4 had no effect of the degradation of IκB-α induced by LPS stimulation. These results indicate that compound 4 mediated inhibition of pro-inflammatory mediators and cytokines probably occurred via inhibition of JNK signal transduction pathways.
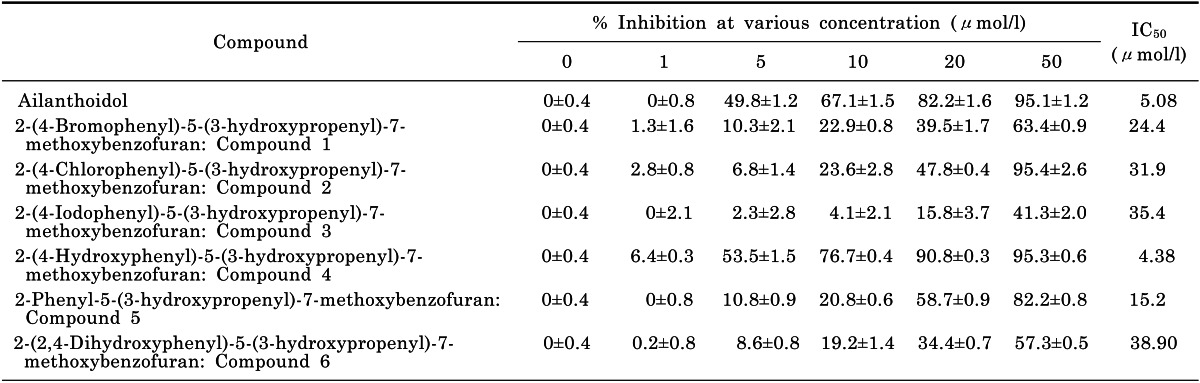
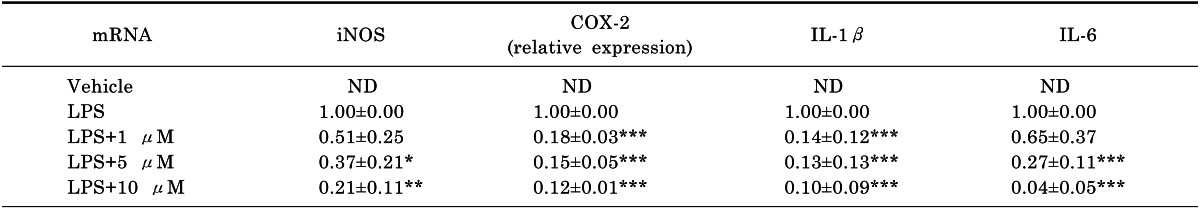
One of the functions of the JNK is the post-translational regulation of AP-1 transcription factors. AP-1 is a dimeric transcription factor composed of proteins with basic leucine zipper domains needed for dimer formation and DNA binding [14]. The major subfamilies that form AP-1 are Jun and Fos. Since the Jun family is activated through the phosphorylation by JNKs, we examined the phosphorylation and nuclear translocation of c-Jun after treatment with compound 4. As shown in Fig. 5B and 5C, compound 4 inhibited the phosphorylation and nuclear translocation of c-Jun induced by LPS stimulation. These results indicate that compound 4 reduced the expression of various inflammatory mediators via inhibition of MAPK (JNK) and AP-1 (c-Jun).
DISCUSSION
Since inflammatory reactions cause various diseases, the discovery of effective and economical new drugs that will control inflammation is important [15]. The discovery of new drugs from natural products including plants and marine products has become a focus of research [2,16-18]. Despite these efforts, several limitations still exist. For example, the active components extracted from natural sources are present in low quantity and their extraction can be complex. To overcome these problems, the use of synthetically manufactured compounds has been explored, based on information gained from active compounds isolated from natural products [13,19-21]. Ailanthoidol is a bioactive component isolated from Zanthoxylum ailanthoides and Salvia miltiorrhiza Bunge [2,13]. Our and other groups developed effective and simple methods to synthesis ailanthoidol [7,8,13], in light of the demonstrations that ailanthoidol has estrogenic [22], antitumor [4] and anti-inflammatory actions [3]. In addition, we constructed several ailanthoidol derivatives to investigate various pharmacological functions [13].
In the present study, we evaluated the anti-inflammatory activities of six ailanthoidol derivatives, compounds 1-6. Among these derivatives, compound 4 significantly inhibited NO release from RAW264.7 cells. Compound 4 also markedly inhibited LPS-induced production of NO and PGE2 through the down-regulation of expression of COX-2 and iNOS in macrophages, respectively, suppressed the expressions of IL-1β and IL-6 at the mRNA and protein levels in macrophages and suppressed LPS-dependent activation of AP-1, but not NF-κB. The results are the first to demonstrate that compound 4 inhibits the inflammatory response in macrophages.
Our previous results showed that ailanthoidol significantly suppresses LPS-induced NF-κB activation, but not AP-1 [3]. In the same study, ailanthoidol was demonstrated capable of inhibiting the LPS-induced degradation of IκB and nuclear translocation of NF-κB. In addition, the DNA binding activity of NF-κB can be blocked by ailanthoidol in LPS-stimulated RAW264.7 cells [3]. Interestingly, compound 4 could not block NF-κB signaling pathway, although its structure is very similar to ailanthoidol. Instead of the NF-κB signaling pathway, phosphorylation of JNK as well as c-Jun was inhibited by compound 4. Although these structural analogs differ at only one position, their effect on NF-κB and AP-1 signaling are quite different. Further studies will be needed to demonstrate the relationship between the structural specifications of ailanthoidol derivatives and their anti-inflammatory molecular mechanism.
The present results provide some preliminary but useful insights into the molecular mechanisms of compound 4. Compound 4 did not affect the degradation of IκB-α, but suppressed the phosphorylation of JNK and c-Jun, as well as nuclear translocation of c-Jun, which were induced by LPS stimulation. To the end of the signaling generated by LPS in macrophages, the activation of transcription factors results in the production of both pro- and anti-inflammatory mediators. The binding of LPS to TLR-4 leads to activation of transcription factor NF-κB and AP-1, which regulate innate immune responses [23]. Activation of NF-κB and AP-1 induces the expression of several inflammatory mediators such as iNOS, COX-2, IL-1β and IL-6, along with many other genes [24,25]. AP-1 is composed of proteins belonging to the Jun and Fos families, and c-Jun and c-Fos are immediate-early genes [14,26]. MAPK signaling pathways regulate AP-1 activity by increasing transcription and the phosphorylation of AP-1 proteins.
These results suggest that the inhibitions of NO, PGE2 and pro-inflammatory cytokines in compound 4 treated RAW264.7 cells are caused by the down-regulation of AP-1 transcription factors through the inhibition of the MAPK signaling pathway. A further challenge is to delineate in vivo actions of compound 4 to provide a better understanding of the health-promoting effects of a synthetic compound that is widely consumed globally.
 XML Download
XML Download