

 PDF
PDF ePub
ePub Citation
Citation Print
Print


ABBREVIATIONS
GVHD
graft-versus-host disease
BM
bone marrow
HSCs
hematopoietic stem cells
NK
natural killer
SCF
stem cell factor
IL
interleukin
FLT3L
fms-like tyrosine kinase-3 ligand
HDC
hydrocortisone
FCS
fetal calf serum
DMEM
Dulbecco's modified eagles medium
FITC
fluorescein isothiocyanate
PE
phycoerythrin
HUVECs
human umbilical vein endothelial cells
SPF
specific pathogen-free
INTRODUCTION
The porcine is a useful animal model for biomedical research in xeno-transplantation as well as in regenerative medicine. Organ size and physiological similarities between humans and porcine make these animals suitable organ transplantation for humans [1,2]. However, organ transplantation across species is prone to vigorous immunologic rejection. Transplantation of whole bone marrow (BM) cells or selected subsets has been a successful strategy to achieve chimerism and induction of tolerance in different xeno-transplantation studies [3,4]. Nevertheless, in spite of promising results, this approach remains limited in part because certain BM cell subpopulations may cause graft-versus-host disease (GVHD).
BM-derived stem cells are the most primitive cells that establish a fully self-renewing population and retain multilineage hematopoietic differentiation potential. Unlike whole BM, stem cell-enriched populations do not contain mature lymphocytes capable of inducing GVHD. In the adult BM, most hematopoietic progenitors express c-kit (CD117), a member of a family of cell-surface receptors with tyrosine kinase activity. Purified c-kit+ BM cells have been used in mice to induce stable multilineage mixed chimerism and permanent acceptance of donorspecific grafts without any clinical signs of GVHD [3]. Human hematopoietic stem cells (HSCs) with long term engrafting abilities are also almost exclusively in the c-kit+ population of CD34+ BM cells [5]. On the other hand, the recent demonstration of the capacity of BM-derived HSCs to differentiate into nonhematopoietic lineages such as hepatocytes, skeletal muscle cells, neurons or epithelial cells of the lung, gastrointestinal tract and skin, has raised hopes as to their therapeutic potential in regenerative medicine [6].
Natural killer (NK) cells are innate immune effector cells. Their derivation from either lymphoid or myeloid lineages was debated early in their discovery [7]. Further research showed that NK cells can be derived from common lymphoid progenitors and hence have been considered separate from myeloid lineage [8,9]. However, some studies question this and have shown that progenitors expressing myeloid antigens can develop into NK cells [10,11]. In previous study, it is shown that NK cell differentiation from HSCs can be studied in vitro [12,13]. This process depends on cytokines, notably interleukin-2 (IL-2) or IL-15, whereas other factors (stem cell factor [SCF], fms-like tyrosine kinase-3 ligand [FLT3L]) induce early HSCs expansion and responsiveness to IL-2 and IL-15 signaling [14]. To date, the factors regulating NK differentiation from porcine c-kit+ BM cells are poorly understood. However, it is certain that it involves cell to cell interactions that render NK responsive to soluble factors from the microenvironment, and several factors such as IL-15 are reported to be involved in differentiation of NK cells from HSCs [15]. Although the intact BM microenvironment is essential for NK differentiation, the some cytokines might be required by early acting including SCF, FLT3L, and IL-7 [16,17] in porcine as did differentiation to NK cells from human HSCs. In the present report, we investigated the role of a various of cytokines in the development of CD2+CD8+ NK cells from porcine c-kit+ BM cells, and analyzed the cytotoxic activities of CD2+CD8+ NK cells.
Until recently, it is well known that NK cells are effective at clearing tumors of epithelial origin as well as metastases from tumors [18]. Grzywacz et al. showed that human myeloid precursors, hematopoietic stem cells, can differentiate into NK cells and the NK cells can kill humanerythroleukemic cells [19]. Moreover, it is reported that NK cells exhibit spontaneous cytotoxicity against MHC class I-deficient target cells, and they participate in the innate immune responses against transformed cells and tumor metastases in vivo [20]. In addition, it is studied that the porcine NK cells after cytokine stimulation lysis humanerythroleukemic cells [21]. Particularly, Sotiriadis et al. reported that porcine NK cells can lysis human carcinoma cells [22].
As mentioned above, the concept that NK cells can be derived from the c-kit+ BM cells advances our understanding of the NK ontogeny and relationship to other blood lineages, creating the potential for therapeutic applications.
METHODS
Animals
We used 15 male porcines (10~12 weeks old, 20~25 kg) purchased from Samtako (South Korea). An automatically controlled 12 h photoperiod (lights on 07:00 a.m. and off 07:00 p.m.) and thermostatically regulated ambient temperature (20~21℃) were maintained throughout the experiment. The porcines were housed individually in stainless steel crates, fed the standard, completely balanced pelleted dry food twice a day (07:00~07:30 a.m. and 02:00~02:30 p.m.), with water available ad libitum. The animals were acclimated to their new environment for about 1 week before the experiments. All efforts were made to minimize animal suffering and to reduce the number of animals used, with all the protocols being reviewed and approved by the Animal Research Ethics Committee of the Chonbuk National University (Jeonju, Korea) for the protection of vertebrate animals used for experimental and other scientific purpose.
Purification of porcine c-kit+ BM cells
Porcines were euthanized and processed into a single cell suspension. Porcine bone marrow cells, containing c-kit+ HSCs, were harvested and isolated by a modification of the technique described previously [23]. Femurs were removed and cleaned of muscle tissue. Femurs were then dissected longitudinally and compressed to excise BM cells. BM cells were collected, washed with Dulbecco's modified Eagles medium (DMEM) containing 1% (v/v) fetal calf serum (FCS) and 5 U/ml heparin, and processed into a single cell suspension. Since then, each sample was diluted in 0.9% sterile saline solution (1:1) (v/v). After dilution was completed, the mixture was added to Ficoll-Paque PLUS (1.077 g/ml; #07907, StemCell Technologies, Vancouver, Canada) in another sterile tube and centrifuged at 400 g for 30 min at room temperature. Using a sterile Pasteur pipette the plasma layer on top was removed and discarded; care being taken to prevent any disturbance of the mononuclear cell layer. And, the aspirate of porcine mononuclear cell layer was collected and transferred into a sterile 17×100 mm polystyrene tube. Ammonium chloride lysis was performed on the remaining pellet to remove red blood cells. Once completely lysed, the pellet was three times with 15 ml of McCoy's 5A culture media, and centrifuged at 400 g for 5 min each time. A cell count was then performed using an automatic cell counter in order to determine the number of nucleated cells in the culture plate. Routine microbiologic monitoring did not reveal any evidence of infection with common pathogens.
Isolation of porcine c-kit+ BM cells
For isolation of c-kit+ BM cells from mononuclear cells, biotinylated porcine SCF was used to detect and purify porcine c-kit+ cells [24]. Briefly, low-density cells separated with Histopaque (Sigma, St. Louis, MO, USA) were incubated for 1 h on ice with biotinylated SCF (320 µg/l×109 BMCs for magnetic bead separation), and then stained for 15 min with streptavidin-coated magnetic microbeads (Miltenyi Biotec, Bergisch-Gladbach, Germany). After extensive phosphate-buffered saline (PBS) washes, c-kit+ cells were sorted by the MACS system (Miltenyi Biotec). Percentages of c-kit+ cells fractions used in this study were greater than 95%.
Developments of cytokines-induced CD2+CD8+ NK cells from porcine c-kit+ BM cells
The human umbilical vein endothelial cells (HUVECs) was cultured to confluence and was irradiated (30 Gy [3000 rad]) [25]. Porcine c-kit+ BM cells were cultured with a monolayer of HUVECs plus Dulbecco modified Eagle medium (DMEM) (1:2 ratio) with 20% porcine serum, ethanolamine (50 µM), ascorbic acid (20 mg/l), 5 µg/l sodium selenite (Na2SeO3), β-mercaptoethanol (24 µM), and penicillin (100 U/ml)/streptomycin (100 U/ml). At the start of cultures, IL-2 (30 ng/ml) , IL-7 (10 ng/ml), IL-15 (10 ng/ml), IL-21 (30 ng/ml), SCF (30 ng/ml), and FLT3L (50 ng/ml) (R&D Systems, Minneapolis, MN, USA) were added. Cultures were refed weekly by 50% volume change of media, supplemented with the above-listed cytokines (except IL-3), with HDC. For 28 days culture, once a week cytokines-activated cells were harvested, counted, and analyzed.
FACS analysis
Phenotypes of the cytokines-induced NK cells were characterized by multi-color flow cytometry analysis (FACS Calibur with two lasers, Becton Dickinson) using combinations of the porcine antibodies to facilitate two color analysis. Cells were stained unfixed in 96 well round bottom microtitre plates (Sterilin, Cambridge, UK). In each case, cells were incubated on ice with the CD2-fluorescein isothiocyanate (FITC) and CD8-phycoerythrin (PE) mAbs for 20 min, washed and further incubated on ice for another 20 min with fluorescent dye conjugated secondary antibody (either isotype specific FITC, Alexa 488, Phycoerythrin, Alexa 633 or Streptavidin-PERCP-Southern Biotechnology Association, Molecular Probes, BD Biosciences). The cells were diluted and washed with FACS diluent; PBS pH 7.4 plus 0.6% bovine serum albumin, 0.1% sodium azide and 2% horse serum. After the external staining the cells were given one final wash and fixed by the addition of 50 ml of FACS-fix (450 ml sterile distilled water plus 60 ml 10× calcium/magnesium free PBS pH 7.4 [Sigma] and 15 ml formaldehyde solution [38% (w/w); Analar]) diluted 1 in 4 with FACS diluent and placed on ice for 1 h. The cells were then washed with FACS diluent and 50 ml of permeabilization solution [FACS diluent plus 0.05% Tween 20 (Sigma)] added to every well. The plates were then sealed and carefully floated on a 37℃ water bath for 15 min. The seal was removed and the cells washed with FACS diluent to remove the detergent.
Reverse transcription and real-time polymerase chain reaction
Total RNA from cytokines-activated cells (5×106 cells/ml) was prepared by adding TRIzol Reagent (Invitrogen, California, USA) according to the manufacturer's protocol [26]. The RNA solution was stored at -70℃ until used. Semiquantitative reverse transcriptase reactions were conducted using MuLV reverse transcriptase. One milligram of RNA was incubated with oligo-dT15 for 5 min at 70℃ and then mixed with a 5× first-strand buffer, 10 mM dNTPs, and 0.1 M DTT. The reaction mixture was further incubated for 5 min at 37℃ and for 60 min after the addition of MuLV reverse transcriptase (2 U). Reactions were terminated after 10 min at 70℃, and total RNA was depleted by adding RNase H. The polymerase chain reaction (PCR) was conducted with the incubation mixture (2 ml cDNA, 4 mM 5' and 3' primers, a 10× buffer [10 mM Tris-HCl, pH 8.3, 50 mM KCl, 0.1% Triton X-100], 250 µM of dNTP, 25 mM of MgCl2, and 1 unit of Taq polymerase [Promega, Wisconsin, USA]). The following incubation conditions were used: a 30 sec denaturation time at 94℃, an annealing time of 30 sec at 55℃, an extension time of 45 sec at 72℃, and a final extension of 5 min at 72℃. For RT-PCR analysis, one microgram of RNA was submitted to reverse transcription with the Molony murine leukemia virus reverse transcriptase (Invitrogen, California, USA). Two microliters of cDNA obtained from each sample were submitted to a qPCR assay using the SYBR green Master mix method (Life technology, Applied Biosystem, California, USA) in the ABO sequence detection system, The primers (Bioneer, Daejeon, Korea) used in this experiment are indicated as following [perforin F 5' - CTG GCT CAC ACG AGG TGA TG - 3'; perforin R 5' - TCA GGT GAG GCA AGC ACT TG - 3'], [granzyme B F 5' - AAC ACC CAG CTG TGT GTA GG - 3'; granzyme B R 5' - TGA GAC TTT GGT GCA GGC TC - 3'] and [NKG2D F 5' -TCC TCC CAG CAT GGA GAT AAG GGA - 3'; NKG2D R 5' - GTT AGG TGA GAG GAT GGA ACC ATC TTC - 3'], designed from published porcine sequences [27]. Each PCR product signal was normalized relative to that of G3PDH obtained from the same RNA.
Preparation of protein lysate and immunoblot analysis
Lysates of cytokines-stimulated c-kit+ BM cells were separated as previously described [28]. Briefly, cytokines-stimulated CD2+CD8+ NK cells (5×106 cells/ml) washed 3 times in cold phosphate-buffered saline with 1 mM sodium orthovanadate and were lysed in lysis buffer (20 mM Tris-HCl, pH 7.4, 2 mM EDTA, 2 mM ethyleneglycotetraacetic acid, 50 mM β-glycerophosphate, 1 mM sodium orthovanadate, 1 mM dithiothreitol, 1% Triton X-100, 10% glycerol, 10 µg/ml aprotinin, 10 µg/ml pepstatin, 1 mM benzimide, and 2 mM PMSF) for 30 min with rotation at 4℃. The lysates were clarified by centrifugation at 16,000 g for 10 min at 4℃ and stored at -20℃ until needed.
Then, whole cell lysates were analyzed by immunoblotting methods as reported previously [29]. Briefly, proteins were separated on 10% sodium dodecyl sulfate-polyacrylamide gels and were transferred by electroblotting to a polyvinylidenedifluoride membrane. Membranes were blocked for 60 min in Tris-buffered saline containing 3% bovine serum albumin, 20 mM NaF, 2 mM EDTA, and 0.2% Tween 20 at room temperature. The membrane was incubated for 60 min with specific primary antibody [1:1,000 for perforin (Mouse IgG2b; clone No: F-1), granzyme B (Mouse IgG1; clone No: 2C5) and NKG2D (Mouse IgG1; clone No: 1D11) and 1:2,000 for β-actin (Mouse IgG1; clone No: C-4)] at 4℃, washed 3 times with the same buffer, and incubated for an additional 60 min with horseradish peroxidase conjugated secondary antibody. The total levels of perforin, granzyme B, NKG2D, and β-actin were visualized by chemiluminescence using Supersignal West Dura Extended Duration Substrate (Pierce, Rockford, IL, USA), and the intensity of the immunoreactive bands was quantified using a densitometer (Sanyo, Tokyo, Japan). Experiments were performed independently five times.
Cytotoxic assay of cytokines-induced porcine CD2+CD8+ NK cells
The cytotoxicity of procine NK cells was quantified using a 51Cr-release assay according to the procedure described previously in [30]. Briefly, in the NK cytotoxicity assay, the cytokines-induced CD2+CD8+ NK cells were used as the effector cells. The CD2+CD8+ NK cells were incubated in 100-mm culture dish containing 7 ml of DMEM with 20% porcine serum. And, the erythroleukemic tumor cell line; K562 cells obtained from ATCC (American Type Culture Collection, Rockville, MD, USA) were used as targets, This line was maintained in a stationary suspension culture in complete medium: RPMI 1640 supplemented with 10% heat-inactivated fetal bovine serum (FBS) (Sigma, St Louis, USA), penicillin (Sigma, St Louis, USA) (100 U/ml) and streptomycin (Sigma, St Louis, USA) (100 mg/ml) in a 5% CO2 humidified incubator (standard culture conditions). Washed in complete medium, the K562 target cells (5×106) were labeled with 100 µCi of Na2 51CrO4 (Radio Chemical Center, Otwock-Świerk, Poland) at 37℃ for 1 h. The labeled target cells were then washed five times using RPMI containing 2% FBS and adjusted to 1×105/ml in complete medium The targets cells (2×104) were cultured in round-bottomed microwells plates (Nun, Roskilde, Denmark) with various concentrations of effector cells (effector: target cells ratios: 100:1 in a total volume of 200 µl) in triplicate, under standard culture conditions for 4 h. Spontaneous 51Cr release wells had target cells plus 100 µl of complete medium and the maximum release wells contained target cells plus 100 µl of complete medium with 5% Triton X-100 (Sigma, St Louis, USA). The assay was terminated at the 4th h by centrifuging the plates (500×g, 5 min). Then 100 µl of supernatant was removed from each well. An isotope count and the percentage of specific lysis (specific 51Cr release) were performed with a gamma counter (Baker System 9120, Allentown, USA) as follows:
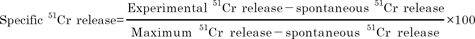
Statistical analysis
All data were expressed as the mean±SD. Data were analyzed for statistical significance using one-way (ANOVA), followed by Tukey's test as a post-hoc test with SPSS software (SPSS for Windows, Version 10). A probability value of p<0.05 was considered to indicate statistical significance, and all experiments were performed at least three times.
RESULTS
Phenotype analysis of porcine c-kit+ BM cells
We examined the expression levels of porcine c-kit, CD117. The fluorescent intensity of Allexa Fluor 488-CD117 was used to measure the expression of c-kit. As shown in Fig. 1A, porcine BM cells were stained with more than 98.9±3.7% compare to isotype control. After 16 h, BM cells were aggregated each other to clump (Fig. 1B).
Cytokines-induced NK cells identified by CD2+CD8+ cells
Surface marker CD2 and CD8 molecules are highly specific for porcine NK cells [31]. Cytokines-activated cells were isolated from co-culture plate with HUVEC. Then, CD2+CD8+ cells were determined using a FACS analysis. Only primary cultured cells were used, because they have the ability to become various types of cells under specific culture conditions. Therefore, cytokines-activated c-kit+ BM cells for 7th, 14th, 21th and 28th days were confirmed by FACS analysis using porcine NK cells marker-specific antibodies (Fig. 2, *p<0.01). The FACS results demonstrate that 1.2±1.7, 5.5±3.4, 9.7±7.3, 35.5±6.8 and 34.7±8.7% of the cytokines-induced NK cells expressed CD2 and CD8 molecules compared to c-kit+ BM cells not pretreated with cytokines (2.5±1.8, 2.7±2.4, 2.8±2.2, 2.2±2.3 and 2.3±1.9%, respectively) in length of time (Fig. 2, *p<0.01). Whereas, all c-kit+ BM cells did not differentiate to CD2+CD8+ NK cells when stromal cells was not used as a feeder cells (data not shown), suggesting stromal cells is necessary for the development of NK cells in porcine.
Analysis of perforin, granzyme B, and NKG2D mRNA expression by RT-PCR
The cytokines-induced CD2+CD8+ NK cells were examined for the perforin, granzyme B, and NKG2D gene expression by RT-PCR. The c-kit+ BM cells not pretreated with cytokines were also examined for the same genes. Fig. 3 shows that the cytokines-stimulated c-kit+ BM cells have a stronger perforin, granzyme B, and NKG2D mRNA expression than the c-kit+ BM cells not pretreated with cytokines in length of time. As shown Fig. 3, perforin, granzyme B, and NKG2D mRNA level were increased for 7th, 14th, 21th, and 28th days compare to day 0 (just before the treatment of cytokines). The perforin and granzyme B mRNA levels have more than about 2-fold 14 days later (Fig. 3A and 3B, *p<0.01). NKG2D mRNA expression was increased about 1.5-fold 21 days later compare to day 0 (Fig. 3C, *p<0.01).
Measurement of perforin, granzyme B, and NKG2D protein levels
Because cytokines may induce NK cell cytotoxicity against cancer cells through a secretion of perforin [32], granzyme B [33], and NKG2D [34] in human or porcine, we examined whether the treatment of cytokines increased perforin, granzyme B, and NKG2D activity using western blotting and fluorescence image. Cytokines-stimulated c-kit+ BM cells were examined for the perforin, granzyme B, and NKG2D protein levels. The c-kit+ BM cells not pretreated with cytokines were also examined for these protein levels in length of time. Fig. 4 shows that cytokines-stimulated c-kit+ BM cells have a stronger perforin, granzyme B, and NKG2D band than the c-kit+ BM cells not pretreated with cytokines in length of time for 28 days. Fig. 4 shows that the cytokines-stimulated c-kit+ BM cells have increased perforin, granzyme B, and FasL levels than the c-kit+ BM cells not pretreated with cytokines in length of time. As shown Fig. 4A and 4B, perforin and granzyme B levels were significantly increased 14 days later treatment of cytokines (*p<0.01). Whereas, NKG2D protein was significantly increased 21 days later treatment of cytokines (Fig. 4C; *p<0.01).
Cytotoxic activities of cytokines-stimulated NK cells
The in vitro cytotoxic property of effector cells (cytokines-induced CD2+CD8+ NK cells) against target cells (K562 cells) was analyzed. The ratios of effector cells and target cells were 100:1. Cytokines-induced CD2+CD8+ NK cells evoked a cytotoxic activity against target cells two weeks later. Cytotoxic activity induced by cytokines was seen in each time-point after cytokines treatment (Fig. 5); 1.28±0.87% (without cytokines) and 1.53±0.52% (with cytokines) at day 0, 1.32±0.76% (without cytokines) and 2.84±0.98% (with cytokines) at 7 day, 1.07±0.98% (without cytokines) and 7.65±2.27% (with cytokines) at day 14, 1.52±0.85% (without cytokines) and 22.76±4.75% (with cytokines) at day 21, and 1.98±0.91% (without cytokines) and 19.53±4.98% (with cytokines) at day 28 (Fig. 5; *p<0.01). A significant increase in cytotoxic activity in response to treatment of cytokines was observed 14 days later treatment of cytokines. The early (before 14 days treatment of cytokines) cytotoxic changes were not significantly difference (p>0.05). These results show that cytokines-induced CD2+CD8+ NK cells could be more cytotoxic activities against target cells than the c-kit+ BM cells not pretreated with cytokines.
DISCUSSION
NK cells are the lymphocytes that mature in the bone marrow, derived from HSC [35]. These NK cells mediate critical functions in innate immunity by directly destroying infected or transformed cells [15]. The factors regulating NK differentiation are poorly understood. However, it is certain that it involves cell to cell interactions that render NK responsive to soluble factors from the microenvironment, and several factors such as IL-15 are reported to be involved in differentiation of NK cells. Although the intact BM microenvironment is essential for NK differentiation, the stromal cells requirements might be replaced, at least partially, by early-acting cytokines including SCF, FLT3L, and IL-7 [16,17]. These cytokines were reported to induce CD122 responsiveness through their receptors in NK cells [15]. Interestingly, despite that IL-15 has been known to be essential for NK generation [36,37], transition from HSC to premature NK does not require IL-15 [38]. Thus the generation of premature NK is regarded as IL-15-independent mechanism. Once committed to the NK lineage, premature NK cells acquire the phenotypic and functional characteristics of mature NK cells by IL-15 stimulation through their receptors. In human, IL-2 can also promote the acquisition of CD19 and KIRs on premature NK or immature NK cells [39], although IL-2 deficient mice exhibit normal population of NK cells, suggesting IL-2 is not necessary for the development of NK cells in mice. Also, in previous reports, it is observed that combination of stromal cells and HDCs may greatly increase the in vitro NK-cell differentiation from CD34+CD38-/+ cells in the presence of appropriate cytokines [10,35,40]. Although much is known about the function of NK cells in humans and mice, NK cells activity in swine remains preliminary, and their role in animal viral diseases still is obscure. The recent progress in these animal species has been reviewed by Boysen and Storset [41] and Gerner et al. [42]. In porcine, NK cells may account for a total of 5 to 10% of circulating lymphocytes and currently are identified as belonging to a subset of cells that coexpress CD2 and CD8 molecules [32]. Although mRNAs related to many cytotoxic molecules have been detected, until now no studies have been conducted to define their role in the generic function of porcine NK cells. But it is known that porcine NK cells can secret perforin, granzyme B and NKG2D, and kill in vitro targets [31-33]. Their function can be modulated by direct stimulation with cytokines such as interleukin-2 (IL-2), IL-12, IL-15, IL-18 [43,44]. Against this background, we selected 6 cytokines (IL-2, IL-7, IL-15, IL-21, SCF, and FLT3L), HDCs, and stromal cells as candidates for factors that induce differentiation of porcine HSCs, based on our hypothesis that such factors also play pivotal roles in the differentiation from porcine HSCs to NK cells. As a results, in present study, we show that a fraction of porcine c-kit+ BM cells can develop into CD2+CD8+ NK cells under the influence of cytokines (IL-2, IL-7, IL-15, IL-21, SCF, and FLT3L), HDCs, and stromal cells. But, there are several other possibilities for the low frequency of CD2 and CD8 molecules, porcine NK cell markers. First, the levels of cytokines required for generation of NK cells from BM cells may have narrow ranges. Second, only a small portion of porcine c-kit+ BM cells despite the right levels of cytokines can acquire NK cell properties. Consistent with this idea, CD2+CD8+ NK cells differentiated from c-kit+ BM cells were a mere about 35.5% (Fig. 2). But, these cytokines-induced CD2+CD8+ NK cells expressed the perforin, granzyme B, and NKG2D mRNA when mRNA levels were analyzed. And, their protein levels were increased more than those in c-kit+ BM cells not pretreated with cytokines (Fig. 3 and Fig. 4), suggesting that the cytokines-induced CD2+CD8+ NK cells possess a mechanisms that express the factors related to NK properties. Therefore, we speculate that these cytokines are required in the stage of NK cells generation from porcine c-kit+ BM cells. Another unsolved question is whether these cytokines we identified play roles completely in differentiation from c-kit+ BM cells to CD2+CD8+ NK cells. Because some cytokines may be released from NK cells during the differentiation, we can also assume that these differentiation are partly involved in the machinery regulated by the earlier differentiated NK cells. Therefore, this study provides the first in vitro evidence of a possible role for a various of cytokines in CD2+CD8+ NK cells development from porcine c-kit+ BM cells. Additionally, in previous study, Sotiriadis et al. described the pig model, specifically germ-free pigs and specific pathogen-free (SPF) mini-pigs, had been used to study various aspects of the immune system, specifically innate immunity [22]. Specifically, it is reported that at least for the first 4 weeks of life, pigs are immunocompetent, owing to complete absence of foreign antigen exposure and subsequent lack of immunoglobulin production [45]. Furthermore, it is known that the porcine molecules are much more similar to those in humans [46]. This allows for study of therapeutic applications in an organism without any xenoreactivity with ADCC or direct cell lysis as well as graft-vs-host disease. Also, this confers a much more immediate pertinence to the use of pigs without immune rejection in human disease. But, although xenograft rejection has been largely resolved, other problems may remain major hurdles. Therefore, we think that the ideal solution to xenograft rejection is development of xenograft specific tolerance. In summary, generation of CD2+CD8+ NK cells from porcine c-kit+ BM cells may present a particularly useful system for destroying malignant cells, not only in the field of veterinary medicine, but also for immunology in general.




 XML Download
XML Download