

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Microglia are resident immune cells in the central nervous system that respond to extracellular stimuli and play a crucial role in host defense and tissue repair in brain [1,2]. However, aberrant activation of microglia has also been proposed to play a pathogenic role in neuro-inflammatory conditions such as ischemia, multiple sclerosis, Parkinson's disease and Alzheimer's disease [3-6]. Under these CNS disease conditions, activated microglia have been suggested to contribute to neurodegenerative process by producing pro-inflammatory mediators such as NO and several cytokines such as tumor necrosis factor-α (TNF-α), interleukin-1β (IL-1β) [7-10]. Therefore, suppression of aberrant microglial activation may have valuable therapeutic potential for the treatment of inflammatory diseases in the CNS caused by activated microglia.
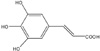
3,4,5-Trihydroxycinnamic acid (THC) is a derivative of hydroxycinnamic acids. Hydroxycinnamic acids have been reported to possess a wide variety of biological properties such as anti-inflammatory, anti-tumor, and neuroprotective activities [11-13]. Ester derivatives of hydroxycinnamic acids have been also reported to exhibit various biological activities [14]. Furthermore, it has been also reported that CAPE, caffeic acid phenylethyl ester, inhibits cytokine-mediated NF-κB activation in murine macrophage cells [15]. Although THC was previously identified in Rooibos tea (Aspalathus linearis) [16], its biological activity has not been extensively examined. Recently, it has been reported that THC inhibits human neutrophil elastase, which causes degradation of healthy tissues contributing to inflammatory diseases such as rheumatoid arthritis and psoriasis [11].
In the present study, in order to provide a novel pharmacological agent that could suppress abnormally activated microglia, the anti-inflammatory activity of THC, a derivative of hydroxycinnamic acids, and its underlying mechanism were examined in LPS-stimulated BV2 microglial cells.
METHODS
Reagents and cell culture
Bacterial lipopolysaccharide (LPS) from Escherichia coli serotype 055:B5 was purchased from Sigma-Aldrich (St. Louis, MO, USA). 3,4,5-Trihydroxycinnamic aicd (THC) (Fig. 1) was purchased from AApin Chemicals Limited (Abingdon, UK). The compound was solubilized in dimethyl sulfoxide (DMSO) and added to the cell culture at the desired concentrations. The BV2 microglial cell line was maintained in Dulbecco's modified Eagle's medium (DMEM; Gibco BRL, Grand Island, NY, USA) containing 5% heat-inactivated fetal bovine serum (FBS; Gibco BRL) and 50 µg/ml gentamicin (Sigma, St. Louis, MO, USA) at 37℃, 5% CO2. In all experiments, cells were incubated in the presence of the indicated concentration of THC before the addition of LPS (200 ng/ml).
Cell viability
Cell viability was determined by 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay. In brief, cells (5×105) were seeded in 12 well plates and treated with various concentrations of THC for 24 hr. MTT (0.5 mg/ml in PBS) was added to each well, and the cells were incubated for 3 hr at 37℃ and 5% CO2. The resulting formazan crystals were dissolved in dimethyl sulfoxied (DMSO). Absorbance was determined at 540 nm. The results were expressed as a percentage of surviving cells over control cells.
Nitrite quantification assay
The production of NO was estimated by measuring the amount of nitrite, a stable metabolite of NO, using the Griess reagent as described [14]. After THC-pretreated BV2 microglial cells were stimulated with LPS in 12-well plates for 24 hr, and then 100 µl of the cell culture media was mixed with an equal volume of Griess reagent. Light absorbance was read at 540 nm. The results were expressed as a percentage of released NO from LPS-stimulated BV2 cells. To prepare a standard curve, sodium nitrite was used to prepare a standard curve.
TNF-α and IL-1β ELISA
The BV2 microglial cells were treated with THC in the absence or presence of LPS. After 24 hr incubation, TNF-α and IL-1β levels in culture media were quantified using monoclonal anti-TNF-α or IL-1β antibodies according to the manufacturer's instruction (R&D Systems).
Western blot analysis
The BV2 microglial cells were pretreated with THC for 1 hr and stimulated with LPS. Cells were washed with PBS and lysed with lysis buffer [50 mM Tris-HCl, pH 8.0, 150 mM NaCl, 0.02% sodium azide, 0.1% sodium dodecyl sulfate (SDS), 1% NP-40, 0.5% sodium deoxycholate, 1 mM phenyl methylsulfonyl fluoride]. Equal amounts of protein were separated on 10% SDS-polyacrylamide gel. Proteins were transferred to Hypond PVDF membrane (Amersham Biosciences, Piscataway, NJ, USA) and blocked in 5% skim milk in TBST for 1 hr at room temperature. Specific antibodies against inducible NO synthase (iNOS), IκB-α (1:1,000; Santa Cruz Biotechnology Inc) and β-actin (1:2,500; Sigma) were diluted in 5% skim milk. After thoroughly washing with TBST, horseradish peroxidase-conjugated secondary antibodies were applied. The blots were developed by the enhanced chemiluminescence detection (Amersham Biosciences).
RNA isolation and Reverse transcription-polymerase chain reaction (RT-PCR)
The BV2 microglial cells were treated with THC in the absence or presence of LPS (200 ng/ml) for 6 h. Total RNA was isolated using the Total RNA Extraction Kit (iNtRON Biotechnology, Inc, USA) according to the manufacturer's instruction. The total RNA (2 µg) obtained from cells was reverse-transcribed using oligo-(dT) 15 primers (Promega, Madison, WI, USA). PCR amplification conditions using primer sets specific for iNOS, TNF-α, IL-1β and β-actin were optimized for each pair of primers. PCR primers were as follows: mouse iNOS forward, 5'-CCCTTC CGAAGTTTCTGGCAGCAG C-3'; iNOS reverse, 5'-GGCTGTCAGAGCCTCGTGGCTTTG G-3'; TNF-α forward, 5'-CATCTTCTCAAAATTCGAGTGACAA-3'; TNF-α reverse, 5'-ACTTGGGC AGATTGACCTCA G-3'; IL-1β forward, 5'-GCAACTGTTCCTGAACTC-3'; IL-1β reverse, 5'-CTCGGAGCCTG TAGTGCA-3'; β-actin forward, 5'-ATCCTGA AAGACCTCTATGC-3'; β-actin reverse, 5'-AACGCAGCTCAGTAACAGTC-3'. Parallel PCR analysis was run for the house keeping gene β-actin to normalize data for differences in mRNA quantity and integrity. PCR products were separated on agarose gel.
Immunofluorescence assay
The BV2 microglial cells were cultured in sterile coverslips and pretreated with THC for 1 hr and stimulated with LPS. After 30 min, the cells were fixed in 4% paraformaldehyde for 20 min at room temperature. The fixed cells were then permeabilized with 0.1% Triton X-100 in PBS and blocked with 3% BSA. Afterwards, the cells were sequentially incubated with rabbit anti-p65 antibody (1 : 100; Santa Cruz Biotechnology Inc) at room temperature and Alexa 546-labeled goat anti-rabbit IgG (Molecular Probes, Eugene, OR, USA) at room temperature for 1 hr. Cells were washed with PBS, and then stained with 0.5 µg/ml of Hoechst staining solution for 10 min at room temperature. After washing with PBS, the coverslips were mounted and observed by confocal microscopy.
Statistical analysis
All values shown in the figures are expressed as the mean±SD obtained from at least three independent experiments. Statistical analysis was carried out by one-way analysis of variance (ANOVA) with Tukey's post-hoc test using SPSS software 12K (SPSS, Chicago, IL, USA). A value of p<0.05 was considered as statistically significant.
RESULTS
THC suppresses NO production in LPS-stimulated BV2 microglial cells
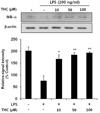
Based on that NO production has been widely used as a representative measure of inflammatory activation of microglia [17], we measured suppressive effects of THC on NO production in LPS-stimulated BV2 microglial cells. Cells were incubated with indicated concentrations of THC (0, 0.1, 1, 10, 25, 50 or 100 µM) for 1 hr prior to LPS treatment and Griess reaction was used as an index for NO synthesis. As shown in Fig. 2A, LPS markedly increased NO production in BV2 microglial cells. However, THC significantly inhibited NO production in LPS-stimulated BV2 micgroglia cells in a concentration dependent manner (Fig. 2A). In addition, THC showed no significant cytotoxicity in concentration ranges, in which THC exhibits suppressive activity of NO production, irrespective of LPS presence (data not shown).
THC inhibits LPS-induced mRNA and protein expression of iNOS
As THC was found to inhibit NO production in Fig. 2A, we examined the mRNA and protein expression levels of iNOS upon THC treatment, using RT-PCR and Western blot analyses, respectively. LPS resulted in markedly increased expression of iNOS protein, which was significantly suppressed with THC treatment in a concentration-dependent manner (Fig. 2B). Consistent with the decrease of iNOS protein, THC also significantly attenuated LPS-induced up-regulation of iNOS mRNA expression (Fig. 2C). These results demonstrate that THC possesses significant inhibitory effects on LPS-induced expression of iNOS, which is considered as a major source of cytotoxic NO.
THC attenuates LPS-induced gene expression and release of pro-inflammatory cytokines
To examine the effects of THC on TNF-α and IL-1β gene expression at the transcriptional level, RT-PCR was performed in the LPS-stimulated BV2 microglial cells. THC significantly suppressed LPS-induced expression of these genes in a concentration-dependent manner (Fig. 3A). In accordance with the change in gene expression of cytokines, THC resulted in a significant suppression of LPS-induced extracellular release of these cytokines (Fig. 3B).
THC attenuates nuclear translocation of NF-κB via suppression of LPS-induced IKB degradation
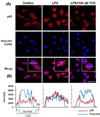
Given the previous report that the transcription factor NF-κB is a predominant regulator of iNOS and numerous pro-inflammatory genes [17,18] and that IκB inhibits the nuclear translocation of p65 subunit of NF-κB, we investigated whether THC inhibits LPS-induced nuclear translocation of NF-κB and degradation of cytosolic IκB. THC blocked LPS-induced degradation of IκB in a concentration-dependent manner (Fig. 4). The cellular localization of p65 was investigated by immunofluorescence staining. Nuclear accumulation of p65 was strongly induced after stimulation of BV2 microglial with LPS (Fig. 5A). However, pretreatment of THC exhibited a significant attenuation of LPS-induced nuclear translocation of p65. Line scanning analysis of confocal images clearly demonstrated the intensity of intracellular localization of p65 subunit (Fig. 5B). These results clearly indicate that THC suppresses LPS-induced NF-κB activation in BV2 microglial cells.
DISCUSSION
The present study clearly demonstrated that THC exhibits anti-inflammatory activity in LPS-stimulated BV2 microglial cells. THC significantly suppressed LPS-induced production of NO, upregulation of iNOS expression, and release of pro-inflammatory cytokines such as TNF-α and IL-1β. Furthermore, THC significantly attenuated LPS-induced IκB degradation and consequent nuclear translocation of NF-κB, a pro-inflammatory transcription factor.
Phenylpropanoids are a group of plant-derived secondary metabolites that are biosynthesized from phenylalanine to coumaric acid and other acids including caffeic acid, ferulic acid, and trihydroxycinnamic acid. Hydroxycinnamic acids and their ester derivatives have been reported to possess a broad spectrum of pharmacological activities including antitumor, antiviral, anti-inflammatory, immunosuppressive, and neuroprotective activities [12-14]. Although it has been, recently, reported that chemically synthesized THC exerts inhibitory activity against human neutrophil elastase, which is involved in the regulation of inflammatory processes [11], the biological properties of THC have not been compared to those of other hydroxycinnamic acids. In the present study, anti-inflammatory activities of THC, originally present in Aspalathus linearis [16], was examined in comparison with caffeic acid, a prototype hydroxycinnamic acid. Both THC and caffeic acid showed significant suppression of LPS-induced NO production in BV2 microglial cells (Fig. 2A). However, THC exhibited significantly greater suppression on LPS-induced NO production compared to caffeic acid. Furthermore, THC showed suppression in a concentration-dependent manner.
Aberrant activation of microglia has been reported to contribute to neuronal damage in several pathologic conditions by releasing pro-inflammatory mediators such as TNF-α, IL-1β and NO [19,20]. Accordingly, suppression of microglial activation and subsequent release of cytokines have been demonstrated to protect neuronal damage in several inflammation-related CNS diseases [3,21,22]. The present study clearly showed that THC suppresses LPS-induced expression of TNF-α and IL-1β and their extracellular release. However, effects of THC on other pro-inflammatory cytokines have not been examined. Therefore, further studies are necessary to examine the effect of THC on the release and gene expression of other cytokines. In addition, the present study showed that THC significantly suppressed LPS-induced NO production, a well-known pro-inflammatory mediator, through the suppression of iNOS gene expression.
NF-κB is an important transcription factor for pro-inflammatory mediators such as iNOS, IL-1β and TNF-α [23,24] and inappropriate regulation of NF-κB and its downstream genes have been associated with various pathological conditions including cancer and autoimmune diseases [25,26]. It has been reported that LPS causes the nuclear translocation of p65 subunit of NF-κB through IκB degradation [27,28]. In accordance with these reports, the present study showed that THC significantly attenuated LPS-induced IκB degradation and consequent nuclear translocation of p65 in LPS-induced BV2 microglial cells (Fig. 5A).
In conclusion, the results clearly demonstrate that THC possesses anti-inflammatory activity such as suppression of NO production and cytokine release by inhibiting nuclear transclocation of NF-κB in LPS-stimulated BV2 microglial cells. The present study strongly suggests that THC might be a valuable therapeutic agent in the treatment of inflammation-related brain pathologies such as ischemia and neurodegenerative diseases. Due to the lack of reports on THC, although no noticeable cytotoxic property of THC was observed in the concentration ranges used in the present study, further studies are necessary to extensively detect any possible toxicities of THC. This endeavor could enhance the possibility of THC for therapeutic application.


 XML Download
XML Download