

 PDF
PDF ePub
ePub Citation
Citation Print
Print


ABBREVIATIONS
TEA
tetraethylammonium
APA
apamin
Glib
glibenclamide
L-NNA
N(G)-nitro-L-arginine
ODQ
1H-(1,2,4) oxadiazolo (4,3-A) quinoxalin-1-one
sGC
an inhibitor of soluble guanylate cyclase
4-AP
4-aminopyridine
KV channel
voltage-dependent K+ channel
NO
nitric oxide
NANC
non-adrenergic non-cholinergic
GI tract
gastrointestinal tract
KBC
K+ channel blockers cocktail
NBC
nerve blockers cocktail
TTX
tetrodotoxin
ATR
atropine
ACh
acetylcholine
SNP
sodium nitroprusside
[Ca2+]i
intracellular Ca2+
VDCCL
voltage-dependent L-type Ca2+ channels
ICC
interstitial cells of cajal
INTRODUCTION
Among various function of stomach [1,2], the motor activity of the proximal part such as receptive (and/or adaptive) relaxation is best known function. Gastric relaxation of proximal stomach from fundus to oral corpus is a key to the accommodation of ingested material. After ingestion, food is largely stored in the proximal stomach and subsequently triturated in the distal stomach. However, full delicate mechanisms are still incomplete, several reflex may be involved [3]. To date, the investigation of gastric reflex has concentrated on the receptive relaxation that the gastric fundus adapts ingested materials [4-6]. In addition to receptive relaxation noted in the process of digestion, relaxation property of proximal stomach is a matter of great interest due to the return of relaxation ability of post operational stomach. Receptive relaxation capacity was usually recovered after subtotal gastrectomy, though it was incomplete. This point suggests that gastric fundus (and/or body) should retain relaxation somewhere and exert its newly adaptive function in a situation requiring relaxation just like after gastrectomy.
The gastric fundus is innervated by excitatory cholinergic neurons and by non-adrenergic non-cholinergic (NANC) inhibitory neurons. NANC inhibitor is thought to be final mediator of the vagally mediated gastric receptive relaxation [7]. Nitric oxide (NO) has been recognized as an inhibitory neurotransmitter to mediate smooth muscle relaxation in the mammalian gastrointestinal (GI) tract [6]. Animal study showed that neurogenic relaxation of gastric fundus is depend on a combination of NO and VIP release [8,9]. Or released NO is believed to be responsible for EFS-induced relaxation [6,10]. In addition, NO was reported to play a central role in gastric fundus relaxations to electrical vagal stimulation in intact animal [6,11] including human [12]. Meanwhile, to data, only a few studies has explored on the nature of neurogenic NANC relaxations elicited either electrically or mechanically in human stomach.
From these reasons, this study was designed to elucidate adaptive relaxation of human gastric corpus using high K+ condition. Several relaxation-related mechanisms were evaluated systematically according to regions (corpus and higher corpus), muscle layers (circular and longitudinal muscle layers), and sides (lesser curvature and greater curvature) in human stomach.
METHODS
Tissue preparation for isometric contraction
Human gastric corpus and higher portion of corpus from both greater and lesser curvature were obtained from patients who underwent gastrectomy. All patients gave written informed consent and this experimental protocol for using human stomach was also approved by the Institutional Review Board for Clinical Research of Chungbuk National University. Specimens from macroscopically normal tissue of neoplastic area were removed immediately after surgical resection of stomach. In Krebs (KRB) solution, specimens were pinned down on sylgard plate. After removal of mucosa and submucosa, muscle strips (0.5×2 cm, 0.5 cm thickness) were prepared from corpus and higher portion of corpus according to muscle direction (circular and longitudinal direction) and mounted to vertical organ bath (25 ml) of isometric contractile measuring system. For confirmation pathologist identified smooth muscles of human stomach using HE staining. In vertical organ bath, one end of strip was tied tightly to the holder and the other side was linked to force transducer by hook type holder (Harvard, USA). Force transducer was connected to PowerLab-Data Acquisition System, which was linked to IBM compatible computer operated by Charter v5.5 software (ADinstruments, Colorado, USA) for measuring isometric contraction. Each strip was stretched passively to resting tension after 1.5~2 hours equilibration. Then contractile responses of the strip to the high K+ (50 mM, 10 min) was repeated three or four times until the responses were reproducible. During this step, high K-induced responses from circular (contraction) and longitudinal (relaxation) smooth muscle of human stomach were reached to maximal level.
Solution and drugs
KRB solution (CO2/bicarbonate-buffered Tyrode) contained (in mM): NaCl 122, KCl 4.7, MgCl2 1, CaCl2 2, NaHCO3 15, KH2PO4 0.93, and glucose 11 (pH 7.3~7.4, bubbled with 5% CO2/95% O2). Equimolar concentration of Na+ was replaced by K+ to make high K+ (50 mM) solution. Sustained tonic contraction by high K+ stimulus in the presence and absence of various blockers in this study was recorded for 10 min and measured at end of recording at 10 min. And pretreatment of various blockers was applied for 10~12 min before the application of high K+. K+ channel blockers cocktail (KBC; was applied (tetraethylammonium, (TEA, 10 mM); apamin (APA, 300 nM); glibenclamide (Glib, 20 µM); Ba2+ (1 mM))) before application of stimulators to block each K+ channel responses. To rule out nerve mediated response, sometimes nerve blockers cocktail (NBC; tetrodotoxin (TTX, 0.4 µM), guanethidine (1 µM), atropine (ATR, 1 µM) was also used. Sometimes phentolamine (Phento, 5 µM) and proparnonol (Propra, 5 µM) was added to NBC for blocking adrenergic response too. All drugs used in this study were purchased from Sigma.
RESULTS
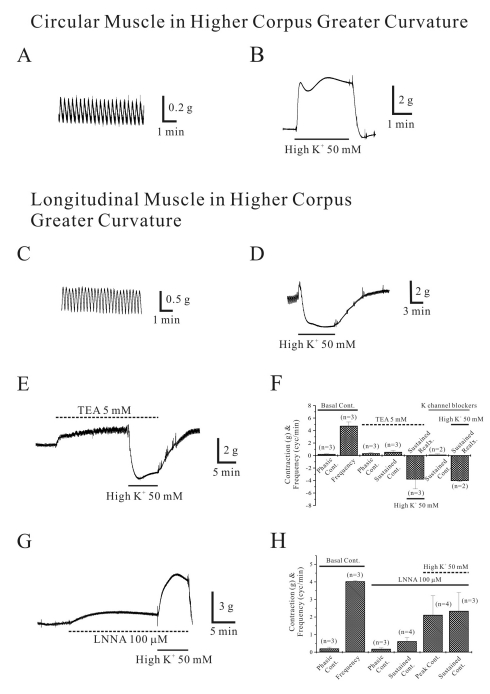
Isometric contraction of circular smooth muscle in corpus greater curvature (GC) of human stomach
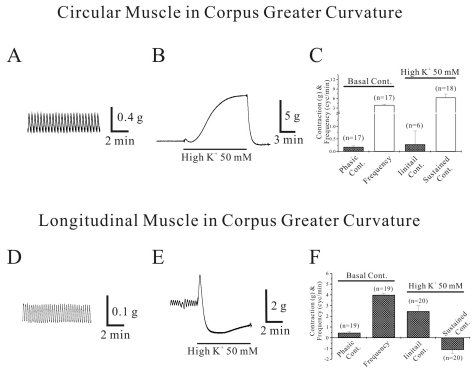
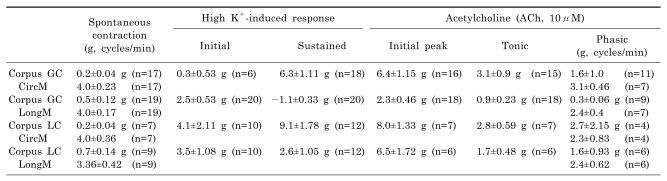
As shown in Fig. 1A, circular smooth muscle from human gastric corpus from GC showed spontaneous contraction of 0.2±0.04 g with a frequency of 4.0±0.23 cycles/min, respectively (n=17, respectively; Fig. 1A and C; Table 1). High K+ produced initial and sustained contraction of 0.3±0.53 g and 6.3±1.11 g, respectively (n=6 and 18, respectively; Fig. 1B and C; Table 1). Acetylcholine (ACh, 10 µM) produced initial transient contraction followed by later sustained tonic contraction and superimposed phasic contractions of 6.4±1.15, 3.1±0.9 and 1.6±1.0 with a frequency of 3.1±0.46 cycles/min, respectively (n=16, 15, 11 and 7, respectively; Table 1). Meanwhile, phasic contraction and its frequency in the absence of ACh was 0.14±0.04 g and 4.8±0.07 cycles/min, respectively (n=7 and 6, respectively; data not shown).
Isometric contraction of longitudinal smooth muscle in corpus GC of human stomach
As shown in Fig. 1D, longitudinal smooth muscle from human gastric corpus from GC showed spontaneous contraction of 0.5±0.12 g with a frequency of 4.0±0.17 cycles/min, respectively (n=19, respectively; Fig. 1D and F; Table 1). However, high K+ produced initial contraction of 2.5±0.53 g followed by sustained relaxation of -1.1±0.33 g (n=20, respectively; Fig. 1E and F; Table 1). High K+-induced relaxation of longitudinal smooth muscle from corpus was not significantly affected by nerve blockers cocktail (NBC, see Methods), too (n=4, p>0.05; bar graph not shown). NBC slightly increased basal tone by 0.15±0.13 g (n=4) and phasic contraction in the absence and presence of NBC were 0.18±1.48 g, 1.0±0.26 cycles/min and 1.20±1.09 g, 2.0±0.82 cycles/min, respectively (n=2, respectively; p>0.05). In the absence of NBC, high K+ produced initial contraction and following sustained relaxation of 2.3±1.73 g and -1.2±0.43 g, respectively (n=4). And the same values in the presence of NBC were 1.2±1.07 g and -1.5±0.32 g, respectively (n=2 and 4, respectively). So, relaxations were not significantly different (p>0.05).
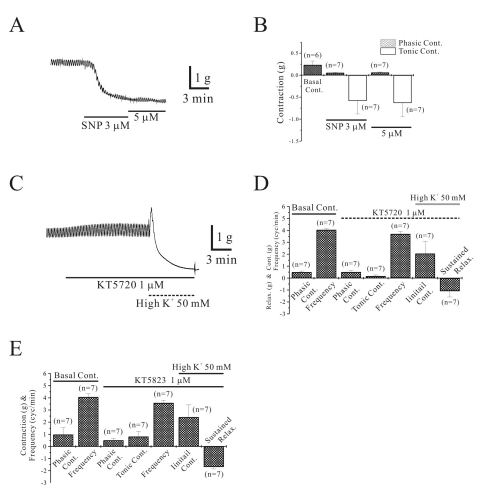
As shown in Fig. 2A and B, we also studied effect of sodium nitroprusside (SNP). SNP (3~5 µM) produced relaxation. In the absence of SNP, phasic contraction was 0.2±0.09 g (n=6) and it was decreased to 0.06±0.01, 0.06±0.01 at concentration of 3 and 5 µM, respectively (n=7 and 7). SNP also inhibited basal tone to -0.58±0.30 and -0.6± 0.31 at concentration of 3 and 5, respectively.
High K+-induced relaxation was also studied in the presence of KT 5720, which is known to inhibit PKA. As shown in Fig. 2C and D, high K+-induced sustained relaxation was not inhibited by pretreatment of KT 5720. In the absence of KT 5720, longitudinal smooth muscle showed phasic contraction of 0.5±0.09 g with a frequency of 4.0±0.10 cycles/ min (n=7, respectively). Application of KT 5720 (1 µM) produced phasic and tonic contraction of 0.5±0.14 g and 0.2±0.09 g with a frequency of 3.7±0.19 cycles/min (n=7, respectively). In the presence of KT 5720 (1 µM), high K+ produced initial contraction and following sustained relaxation of 2.1±1.03 g and -1.1±0.49 g, respectively (n=7, respectively; Fig. 2 and D). The effect of KT 5823, inhibitor of PKG, on high K+-induced relaxation was also studied and it did not affect high K+-induced relaxation (Fig. 2E). In the absence of KT 5823, longitudinal smooth muscle showed phasic contraction of 1.0±0.56 g with a frequency of 4.1±0.27 cycles/min (n=7, respectively). Application of KT 5823 (1 µM) produced phasic and tonic contraction of 0.5±0.15 g and 0.8±0.43 g with a frequency of 3.6±0.21 cycles/min, respectively (n=7, respectively). In the presence of KT 5823 (1 µM), high K+ produced initial contraction and following sustained relaxation of 2.4±1.00 g and -1.7±0.17 g, respectively (n=7, respectively; Fig. 2E).
Effect of L-NNA on high K+-induced relaxation in longitudinal smooth muscle of corpus GC of human stomach
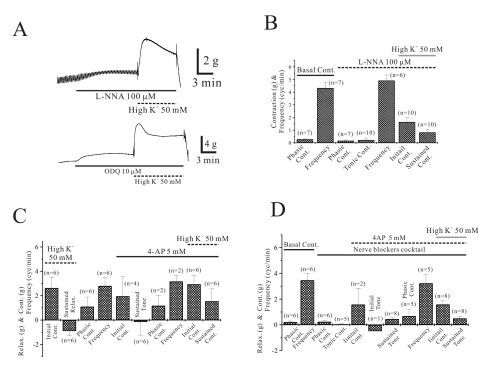
High K+-induced relaxation was also studied in the presence of L-NNA (Fig. 3A and B). In the absence of L-NNA, phasic contraction and its frequency were 0.3±0.07 g and 4.3±0.45 cycles/min, respectively (n=7). As shown in Fig. 3A and B, tonic contraction of 0.2±0.10 g with superimposed phasic contraction of 0.2±0.06 g with a frequency of 4.9±0.44 cycles/min was produced by application of L-NNA (100 µM). And high K+ produced initial and sustained tonic contraction of 1.7±0.34 g and 0.8±0.24 g in the presence of L-NNA (n=10).
High K+-induced relaxation was also studied using 10 µM of ODQ (Fig. 3A). In the absence of ODQ, longitudinal smooth muscle from human gastric corpus showed phasic contraction of 0.7±0.25 g with a frequency of 4.1±0.70 cycles/min, respectively (n=3, respectively). As shown in lower pannel of Fig. 3A, ODQ (10 µM) produced sustained contractions of 0.6±0.31 g (n=5). But high K+-induced relaxation was inhibited in similar fashion with L-NNA as seen in followings. In control, high K+ produced initial contraction and sustained relaxation of 0.9±0.18 g and -1.4±0.54 g, respectively (n=3 and 5, respectively). In the presence of ODQ (10 µM), high K+ produced initial and sustained contractions of 5.0±1.05 g and 2.5±0.73 g, respectively (n=5, respectively; bar graph not shown).
Meanwhile, ACh (10 µM) produced initial transient contraction of 2.3±0.46 g followed by later sustained tonic contraction of 0.9±0.23 g with superimposed phasic contractions of 0.3±0.06 g with a frequency of 2.4±0.4 cycles/min (n=18, 18, 9 and 7, respectively; Table 1). In control, it showed phasic contraction of 0.3±0.06 g with a frequency of 3.9±0.5 g (n=8).
Effects of 4-aminopyridine (4-AP) on high K+-induced relaxation of longitudinal smooth muscle in human gastric corpus GC
High K+-induced sustained relaxation was blocked and reversed to tonic contraction in the presence of 4-AP (Fig. 3C and D). Longitudinal smooth muscle in the absence of 4-AP showed phasic contraction of 1.4±1.01 g with a frequency of 2.7±0.60 cycles/min (n=6; bar graph not shown). And high K+ produced initial contraction and following relaxation of 2.6±0.88 g and -0.9±0.39 g, respectively (n=6, respectively; Fig. 3C). Spontaneous contraction before application of 4-AP again were 1.1±0.80 g and 2.8±0.68 cycles/min (n=6, respectively). And application of 4-AP produced initial contraction of 2.0±1.61 g and then sustained tonic contraction of -0.15±1.09 g with phasic contractions of 1.2±0.85 g, 3.2±0.5 cycles/min (n=4, 6, 2 and 2, respectively; Fig. 3C). In this condition high K+ produced initial contraction of 3.0±0.74 (n=6) and then sustained contraction of 1.5±1.02 g (n=6), instead of relaxation. Therefore, it might imply that high K+-induced relaxation is at least partly mediated by activation of KV channel too.
To check if activation of KV channel is caused by neurotransmitters from longitudinal muscle layer, we studied effect of 4-AP in the presence of NBC (Fig. 3D). In the absence of NBC, it showed phasic contraction of 0.2±0.08 g with a frequency of 3.5±0.5 cycles/min, respectively (n=6, respectively). After application of NBC, it produced sustained tonic and phasic contractions of 0.04±0.06 g and 0.24±0.1 g, respectively (n=6 and 5, respectively; Fig. 3D). Addition of 4-AP produced sustained tonic and phasic contractions of 0.5±1.3 g and 0.7±0.54 g with 3.2±0.69 cycles/min, respectively (n=8, 5 and 5, respectively). In the presence of NBC and 4-AP, high K+ produced initial peak and sustained contractions of 1.6±0.3 g and 0.5±0.17 g, respectively (n=8, respectively; Fig. 3D; p<0.05).
Isometric contraction of circular smooth muscle of corpus lesser curvature (LC) of human stomach
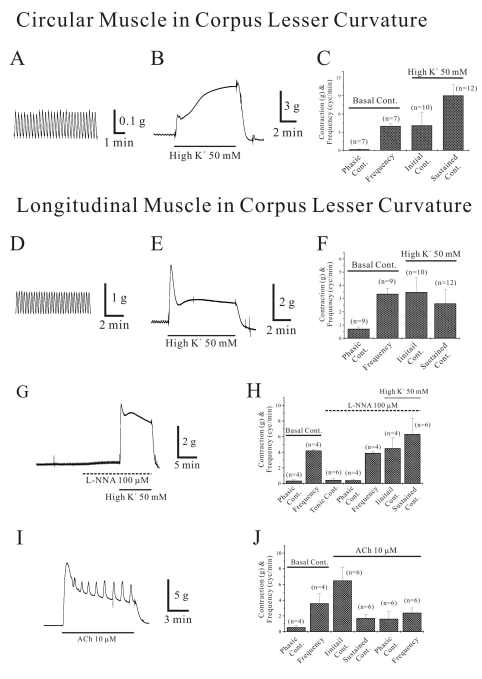
As shown in Fig. 4A and C, circular smooth muscle of human gastric corpus from LC showed spontaneous contraction of 0.2±0.04 g with a frequency of 4.0±0.36 cycles/min (n=7; Table 1). High K+ produced initial and sustained contraction of 4.1±2.1 g and 9.1±1.78 g, respectively (n=10 and 12, respectively; Fig. 4B and C; Table 1). Meanwhile, ACh (10 µM) produced initial transient contraction of 8.0±1.33 g followed by later sustained tonic contraction of 2.8±0.59 g with superimposed phasic contractions of 2.7±2.15 g (n=7, 7 and 4, respectively; Table 1). The frequency of ACh-induced phasic contraction was 2.3±0.83 cycles/min (n=4; Table 1). In control, its phasic contraction and frequency were 0.1±0.02 g and 4.4±0.37 cycles/min, respectively (n=5).
Isometric contraction of longitudinal smooth muscle of corpus LC of human stomach
As shown in Fig. 4D and F, longitudinal smooth muscle from human gastric corpus from LC showed spontaneous contraction of 0.7±0.14 g with a frequency of 3.36±0.42 cycles/min, respectively (n=9, respectively; Table 1). High K+ produced initial and sustained contraction of 3.5±1.08 g and 2.6±1.05 g, respectively (n=10 and 12, respectively; Fig. 4E and F). We also studied effect of L-NNA on high K+-induced contraction. In control, longitudinal smooth muscle from LC produced phasic contraction of 0.4±0.1 g with a frequency of 4.2±0.13 cycles/min (n=4; Fig. 4H). When L-NNA (100 µM) was applied, it produced tonic and phasic contractions of 0.5±0.16 g and 0.4±0.11 g, respectively with a frequency of 3.9±0.15 cycles/min (n=6, 4 and 4, respectively; Fig. 4G and H). In the presence of L-NNA, high K+ produced initial and sustained contraction of 4.5±1.4 g and 6.3±2.0 g, respectively (n=4 and 6, respectively; Fig. 4G and H). Meanwhile, ACh (10 µM) also produced initial transient contraction of 6.5±1.72 g followed by later sustained tonic contraction of 1.7±0.48 g with superimposed phasic contractions of 1.6±0.93 g (n=6; Fig. 4I and J; Table 1). The frequency of ACh-induced phasic contraction was 2.4±0.62 cycles/min (n=6). In control, phasic contraction and its frequency were 0.6±0.20 g and 3.6±1.22 cycles/min, respectively (n=4; Fig. 4J).
Isometric contraction of circular and longitudinal smooth muscle in higher portion of corpus GC of human stomach
Circular and longitudinal smooth muscle showed spontaneous contractions of 0.1±0.03 g with frequency of 3.82±0.31 cycles/min (n=5) and 0.4±0.12 g with frequency of 3.4±0.26 cycles/min (n=4; Fig. 5A, C, D and F), respectively. High K+ produced initial peak contraction of 2.6±1.98 g (n=5) and sustained tonic contraction of 5.8±1.78 g (n=6) (Fig. 5B) in circular muscle. However, high K+ produced initial contraction of 1.6±0.53 g and following sustained relaxation of -1.94±0.99 g in longitudinal muscle (n=6; 5D). Treatment of TEA 5 mM produced phasic contraction of 0.4±0.15 g and tonic contraction of 0.6±0.27 g from basal tone of 0.2±0.04 g with a frequency of 4.8±0.6 cycles/min (n=3). In the presence of TEA, High K+ produced sustained relaxation of -3.8±1.53 g (n=3, Fig. 5E and F). High K+-induced relaxation was not affected by KBC (see Methods) to produce sustained relaxation of -4.1±0.12 g (n=2; Fig. 5F), though minimal tonic contraction of 0.2±0.1 g was produced in two cases. However, high K+-induced relaxation was blocked by pretreatment of L-NNA (Fig. 5G and H). Pretreatment of L-NNA (100 µM) produced phasic contractions of 0.2±0.06 g and sustained tonic contraction of 0.6±0.21 g from basal tone of 0.2±0.08 g with a frequency of 4.0±0.03 cycles/min (n=3). And instead of relaxation high K+ produced peak and sustained contractions of 2.1±1.09 g and 2.4±1.05 g, respectively (n=4 and 3, respectively).
DISCUSSION
In the present study, we found high K+ stimulation produced sustained relaxation in longitudinal muscle from GC of human gastric corpus and higher portion of corpus. Since high K+ (50 mM)-induced relaxation was found in longitudinal muscle not circular muscle in GC side only, this relaxation seem to be regional and muscle specific phenomenon. Furthermore, we observed high K+ (50 mM)-induced relaxation was inhibited by L-NNA, ODQ and KV channel blocker. Therefore, these results suggest that longitudinal muscle from GC of gastric corpus and higher portion of corpus might produce high K+-induced relaxation via activation of NO/sGC pathway and by KV channel.
One of our concerns of this experiment is recovery of relaxation ability of proximal stomach after gastrectomy. We observed this relaxation was highly enhanced in patient who underwent repetitive gastrectomy (in submission). Direct complications of gastrectomy are loss of gastric reservoir, which reduces volume of food intake, and loss of grinding activity. Interestingly, gastric capacity after subtotal gastrectomy could be recovered by proper alimentotherapy although recovery was not fully recovered. Since subtotal gastrectomy saves proximal part of stomach, its ability of relaxation is very important which will be limiting factor for the recovery of gastric function. At present, regional differences and muscle specific gastric relaxation and/or its mechanism in human stomach were not evaluated yet. Furthermore, this relaxation was not observed in circular muscle and not in longitudinal muscle from lesser curvature, either.
To date, both circular and longitudinal muscle is known to be regulated by slow waves [13]. However, we found dissociative coordination between circular and longitudinal muscle of human stomach. This dissociation could be interpreted in a harmonious way that when Ca2+ influxes into muscle cell by high K+ stimulation circular muscles contract and longitudinal muscles relax. To date, the mechanism underlying each muscle layer's different activities is unclear: marked differences of intracellular Ca2+ ([Ca2+]i) regulation by K+-depolarization [14-16]. Dissociative patterns of motility between circular and longitudinal muscle might came not only from the neural activity but also from the different properties of the muscles [14].
In general, Ca2+ is essential for contraction of visceral smooth muscles and it can be assumed that the amplitude of the contraction depends on [Ca2+]i [17]. To date, voltage-dependent 'L-type' Ca2+ channels (VDCCL) have been reported in most excitable tissues, including human stomach [18,19], and VDCCL are known to play a essential role in the regulation of [Ca2+]i in smooth muscles [20]. And the spontaneous contractile activity of GI muscles such as peristaltic contraction is closely related to the slow wave, and the activation of plateau Ca2+ current on it is necessary for the excitation-contraction coupling [21,22]. Therefore, the investigation of regulation of VDCCL is important for understanding gastric functions. In this study, we studied effect of high K+-induced contraction in human stomach (Fig. 1B, C, 4B, C, 4E, F and 5B). High K+ produced contraction activated by VDCCL in gastric circular muscles [14]. Generally, high K+ Stimulation is known to provoke contraction via depolarization and activation of VDCCL [14]. In this study, however, we observed unexpected high K+-induced relaxation in longitudinal muscle of corpus and higher portion of corpus from GC side. Relaxation of longitudinal smooth muscle in contrast to circular muscle might be produced in various reasons; 1) the dissociation of motility and regulation of [Ca2+]i 2) different properties of the muscle fibers 3) different spontaneous activity in each muscle [14,16]. As described in the result, high K+-induced relaxation of corpus was not affected by pretreatment of NBC (p>0.05). Although data not shown, high K+-induced relaxation in corpus region was not also affected by KBC treatment as in the case of higher portion of corpus (Fig. 5E and F). Furthermore, since this relaxation was significantly suppressed by 4-AP with/without NBC, different characteristics of NO pathway and KV channel between circular and longitudinal muscles might be one explanation. Otherwise, unknown difference of cytoskeleton and/or its regulation by mechanical stimulation might underlie this relaxation since stretch-dependent activation of Ca2+ influx via activation of microfilament in gastric circular muscle was reported [23,24].
Gastric accommodation is explained in various mechanisms. In particular, NO are principal candidates for fundus relaxation [25]. In accordance with this fact we found the L-NNA inhibited high K+-induced relaxation in longitudinal smooth muscle of corpus/higher corpus GC (Fig. 3A, B, 5G and H). Furthermore, ODQ (10 µM) blocked high K+-induced relaxation (Fig. 3A and B). In Fig. 2C~E, we also studied involvement of protein kinase G on high K+- induced relaxation since NO is known to activate guanylate cyclase. However, PKG (KT5823) including PKA inhibitor (KT5720) did not affect it. Interestingly as shown in Fig. 1B, high K+-induced contraction of some circular muscle is also followed by small transient relaxation, which was blocked by L-NNA. Thus, this is the first report that NO and/or ODQ directly mediated K+-induced relaxation of longitudinal muscle in human stomach.
NO are reported localized in the majority of myenteric neurons including human gastric fundus and are released to produce proximal gastric relaxation [12]. In addition, it was reported that relaxation response to graded mechanical distention was mostly ascribed to passive viscoelastic properties, with a slight NO-mediated neurogenic component. Though there are still confusions on regulatory mechanisms, which seem to be different depending on how to be stimulated, NO was reported to initiate proximal gastric accommodation in human stomach [12]. Our data point that high K+-induced relaxation was totally blocked by L-NNA and was, moreover, transited to sustained contraction in longitudinal muscle (Fig. 3A, B, 5G and H). This observation strongly indicates that NO is responsible for high K+-induced relaxation and the effect of NO override sustained contraction in human corpus longitudinal muscle [26].
As shown in Fig. 3 and 2C~E, high K+-induced relaxation of longitudinal muscle was blocked and transited to contraction by 4-AP and ODQ but not by KT5823 and KT5720. These observations mean that NO pathway is involved but its downstream signal pathways were not responsible for this relaxation. In smooth muscle, KV channel [27, 28] and KCa channel [29] were reported to be directly regulated by NO/sGC pathways. And nNOS neurons was also found in myenteric border of human stomach [30]. In fact, interstitial cells of Cajal (ICC) expressed NO-sensitive guanylate cyclase and is primary target of NO released from nNOS neurons. And this NO signals are transmitted to smooth muscle in GI tract [31,32]. From these findings we could conclude that high K+ produced muscle relaxation by activation of VDCCL, which was ascribed to NO/sGC-dependent pathways of ICC in longitudinal muscle layer.
In this study, we observed NO-mediated relaxation in longitudinal muscle. And high K+-induced relaxation was not affected by NBC itself in corpus longitudinal muscle too (p>0.05). In fact, NOS neurons are found in circular and longitudinal muscle layers including myenteric border of GI tract [30]. However, NO produced relaxation in longitudinal smooth muscle of human gastric GC side only. Unfortunately, however, we could not exactly explain where NO originate, why longitudinal muscles only produce relaxation by high K+ stimulation in GC side. However, some ICC might be involved in this phenomenon since this relaxation was not affected by NBC [33-36]. Therefore, we will try to find some different characteristics of circular and longitudinal muscle and NO origin in following future.
 XML Download
XML Download