

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Numerous musculoskeletal disorders such as carpal tunnel syndrome, Achilles tendinopathy, and adhesive capsulitis are caused by thickened ligament, tendon stiffness, or fibrosis of the joint capsule. Entrapment neuropathy has been suggested to be caused by increasing compartment pressure due to stiff and thickened ligament structures surrounding the nerve. Increased pressure on a nerve can compress the neural microvasculature and alter blood flow dynamics. High pressures can lead to epineurial arterial ischemia and impaired venous outflow, resulting in venous stasis. This can cause capillary leakage, intraneural edema, or extraneural edema. Consequently, chronic compression can result in inflammation, fibrosis, demyelination, and ultimately axonal loss (1). Pathophysiologic characteristics of adhesive capsulitis include fibrotic tissue changes due to decreased collagen length and fibrofatty infiltration into capsular recess (2). The usual treatments for those conditions include local steroid injections, physical therapy, and administration of nonsteroidal anti-inflammatory drugs. However, these treatments do not decrease compartment pressure or ligament stiffness. They can only provide symptomatic relief. If conservative treatments are ineffective, surgical treatment may be necessary (23).
Relaxin, a peptide hormone, can exert collagenolytic effect on ligamentous and fibrotic tissues (4). In 2002, Hsu and colleagues (5) reported that orphan G-protein receptors LGR7 and LGR8 were relaxin receptors. LGR7 and LGR8 are now known as relaxin family peptide receptors 1 (RXFP1) and 2 (RXFP2), respectively (6). It has been demonstrated that relaxin can bind and activate both RXFP1 and RXFP2 in in vitro cell models (7).
Because relaxin can decrease compartment pressure and relax ligament, tendon, and fibrotic tissue, we hypothesized that relaxin could be used to treat local entrapment neuropathy, tendon stiffness, and adhesive capsulitis. Since hormonal effect depends on the receptor of the hormone at target cells, it is important to confirm the presence of hormonal receptor at target tissues. The effect of relaxin on ligament tissue of knee has been described in ovariectomized adult female rats (8). However, there is limited research on the presence or the distribution of relaxin receptors in various ligaments, tendons, or fibrous tissues of young male Wistar rats. Therefore, the objective of this study was to determine whether relaxin receptors were present in the ligaments, Achilles tendons, or shoulder capsules of young male Wistar rats and to identify the distribution of relaxin receptors in these issues.
MATERIALS AND METHODS
Animals and biological samples
Six 120-day-old male Wistar rats with weight of 180-220 g were obtained from Oriental-Bio Co. (Seoul, Korea). They were euthanized with anesthetic overdose to obtain transverse carpal ligaments, inguinal ligaments, patellar ligaments, anterior cruciate ligaments (ACLs), Achilles tendons, and shoulder joint capsules. Western blot analysis was used to identify relaxin receptor isoforms RXFP1 and RXFP2. The distribution of RXFP1 or RXFP2 in those tissues was determined by immunohistochemical staining.
Protein expression using western blot analysis
After removing the bony attachment to the left forearm and left leg of each rat, tissues described above were snap-frozen in liquid nitrogen and stored at -80°C until analysis. Protein was extracted from 50 mg of tissue (wet weight) using PRO-PREP (Intron Biotech., Seongnam, Korea). An equal amount of protein from each tissue lysate was mixed with a loading dye, boiled for 5 minutes, and subjected to sodium dodecylsulfate-polyacrylamide gel electrophoresis, 10%. Separated proteins were then transferred onto polyvinylidene fluoride membrane (Bio-Rad, Hercules, CA, USA) and blocked with 5% nonfat milk overnight at 4°C. Rabbit anti-Insig1 (RXFP1) polyclonal antibody (Bioss, GA, USA) and rabbit anti-GPR106 (RXFP2) polyclonal antibody (Bioss) were used as primary antibodies with common reactivity to rat and mouse. The membrane was then incubated with rabbit anti-Insig1 (RXFP1) polyclonal antibody (1:200, Bioss), rabbit anti-GPR106 (RXFP2) polyclonal antibody (1:200, Bioss), and rabbit monoclonal anti-glyceraldehyde-3-phosphate dehydrogenase (GAPDH) antibody (1: 1,000, Cell Signaling Technology, Inc., Danvers, MA, USA) diluted in phosphate-buffered saline (PBS) (Sigma-Aldrich, Ontario, Canada) containing 1% bovine serum albumin and Tween-20 (Sigma-Aldrich) and incubated at 4°C for 2 hours. Blots were then washed 3 times for 5 minutes each time and incubated with horseradish peroxidase conjugated anti-rabbit IgG secondary antibodies (1:2,000, Santa Cruz, TX, USA) at room temperature for 1 hour. The membrane was then washed and examined with Pico EPD Western blot detection kit (ELPIS biotech. Inc., Daejeon, Korea) to enable visualization of protein bands. Photographs of blots were captured with a ChemiDocTM MP system (Bio-Rad). The density of each band was determined with Image LabTM Software (Bio-Rad). The ratio of each target-adjusted intensity of band was calculated and considered as the relative expression level of target proteins.
Immunohistochemistry
After removing the bony attachment of the forearm and right leg of each rat, each tissue sample was removed and frozen in liquid nitrogen. Sections (8 μm each) of these frozen tissues were stored in a cryostat at -20°C and transferred to silanized slides, kept at room temperature for 30 minutes, fixed in 4% paraformaldehyde buffered with PBS for 20 minutes, incubated in 0.1 M glycine for 5 minutes, and treated with 0.3% hydrogen peroxide for 10 minutes to remove endogenous peroxidase activity. These sections were then incubated in blocking buffer (0.01% saponin and 3% albumin in PBS) and placed in a humidified chamber at room temperature for 1 hour. Endogenous biotin was blocked with Avidin/Biotin Blocking Kit (Vector Laboratories, Burlingame, CA, USA).
The sections were incubated with primary antibodies diluted at 1:100 in blocking buffer at 4°C overnight. Rabbit anti-Insig1 polyclonal antibody (Bioss) was used as the primary antibody against RXFP1. Rabbit anti-GPR106 polyclonal antibody (Bioss) was used as the primary antibody against RXFP2. Both of these antibodies could recognize rat RXFP proteins.
These sections were rinsed 3 times for 5 minutes each time with blocking buffer and incubated for 1 hour with goat anti-rabbit IgG secondary antibody (1:300, Thermo Fisher Scientific Inc., Rockford, IL, USA). These sections were then rinsed again 3 times for 30 minutes each time with blocking buffer and incubated at room temperature with avidin-biotin-peroxidase complex (Vector Laboratory, Burlingame, CA, USA) for 1.5 hours. Peroxidase activity was revealed by incubation the sections with 3, 3'-diaminobenzidine (0.5 mg/mL) in 0.01% H2O2 for 3 minutes. These sections were then washed and counterstained with hematoxylin using standard protocols. Photographs were taken with Q-imaging Retig 200R camera connected to a Nikon Eclipse 50i microscope (Nikon, Melville, NY, USA).
RESULTS
RXFP1 and RXFP2 proteins expression in each tissue
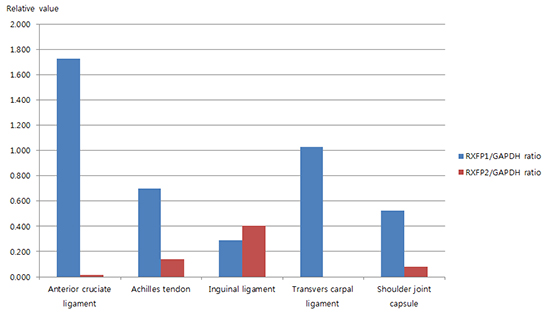
The intensity of RXFP1/GAPDH ratio in each tissue samples is summarized in Table 1. RXFP1 protein was detected in all tissues examined. The protein level of RXFP1 was the highest in ACL sample, followed by transverse carpal ligament, Achilles tendon, shoulder joint capsule, and inguinal ligament samples (Fig. 1). The intensity of RXFP1/GAPDH ratio in each tissue sample is shown in Table 2. RXFP2 protein was not detected in transverse carpal ligament samples. However, it was identified in other samples collected. Its level was the highest in inguinal ligament samples, followed by Achilles tendon, the shoulder joint capsule, and the ACL samples (Fig. 2).
Table 1
The ratio of RXFP1 to GAPDH
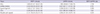
Fig. 1
Ratio of RXFP1 to GAPDH protein in tissue samples studied.
GAPDH, glyceraldehyde-3-phosphate dehydrogenase.

Table 2
The ratio of RXFP2 to GAPDH


Immunolocalization of RXFP1 and RXFP2 proteins in each tissue
Immunohistochemical analyses were performed to reveal the distribution of RXFP1 or RXFP2 in each tissue sample. RXFP1 was identified in vascular endothelial cells in the inguinal ligament samples. It was also found in fibroblast-like cells of ACL and shoulder joint capsule samples as well as in mixed cells of the Achilles tendon and transverse carpal ligament samples (Fig. 3A-3E). RXFP2 was detected in vascular endothelial cells and fibroblast-like cells of the Achilles tendon samples. It was also found in fibroblast-like cells of the inguinal ligament, shoulder joint capsule, and ACL samples (Fig. 4A-4E). In shoulder joint capsule and ACL samples, immunostaining revealed that relaxin receptors were mainly distributed in fibroblast-like cells.
Fig. 3
Immunolocalization of RXFP1 proteins in rats. (A) Anterior cruciate ligament (× 40), (B) Achilles tendon (× 20), (C) Inguinal ligament (× 10), (D) Transverse carpal ligament (× 20), (E) Shoulder joint capsule (× 20). Positively stained vascular endothelial cells are shown in white arrow. Positively stained fibroblast-like cells are shown in black arrow.
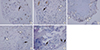
Fig. 4
Immunolocalization of RXFP2 proteins in rats. (A) Anterior cruciate ligament (× 40), (B) Achilles tendon (× 20), (C) Inguinal ligament (× 10), (D) Transverse carpal ligament (× 20), (E) Shoulder joint capsule (× 20). Positively stained vascular endothelial cells are shown in white arrow. Positively stained fibroblast-like cells are shown in black arrow.
DISCUSSION
The effect of relaxin on ligament laxity has been previously reported. In 1926, Hisaw found an aqueous extract of the corpora lutea of pregnant sows caused by interpubic ligament formation in estrogen-primed guinea pigs (9). The hormone contained in that extract that relaxed symphysis pubis was called "relaxin". The relaxin subfamily consists of the following seven peptides: relaxin-1, relaxin-2, relaxin-3, and 4 other insulin-like peptides (INSL3, INSL4, INSL5, and INSL6). They are polypeptide hormones that possess structural similarity to insulin (10). The relaxin family peptides possess their physiologic effect by activating a group of 4 G-protein–coupled receptors (GPCRs), i.e., the relaxin family peptide receptors 1-4 (RXFP1-4). Human relaxin H2 and INSL3 are leucine-rich repeat-containing GPCRs. They are cognate ligands for RXFP1 and RXFP2, respectively (11). Human relaxins H1 and H2 can activate both RXFP1 and RXFP2 while rat relaxin-1 can only weakly bind to RXFP2. RXFP1 and RXFP2 are the two main relaxin receptor isoforms (12). Relaxin-3/RXFP3 system is thought to function primarily in mouse brain (13) while INSL5/RXFP4 system stimulates appetite and activates colon motility to control food intake and glucose homoeostasis in mouse intestine (14).
The primary focus of our study is on RXFP1 and RXFP2 receptors associated with ligaments, tendons, and fibrotic joint capsules. A few reports have revealed the association between ligaments and relaxin receptors. In humans, relaxin receptors have been identified in ACL of men and women, with higher expression in women. This gender-specific difference suggests that estrogen priming might increase the release of endogenous relaxin, the response of target organs to relaxin, and the number of relaxin-binding sites on human smooth muscle or rat myometrium (15). Lubahn and colleagues have reported that relaxin can bind to the volar oblique ligament with specificity (16), suggesting the presence of either cellular or extracellular matrix receptors and a receptor-mediated process. Relaxin receptors are also found in the ligaments, cartilage, and synovium of temporomandibular joint, indicating that these sites might be potential targets for relaxin therapy (17). Recently, the expression of relaxin receptor RXFP1 and RXFP2 has been shown to be regulated by sex steroids in the ligaments and tendons of ovariectomized adult female rat knee (8). However, the studies that yielded those results were restricted to the association of ligaments and joint instability. To further expand those findings, we determined whether relaxin receptors were present in shoulder joint capsules or the tendinous and ligamentous tissues affected by musculoskeletal disorders. Because the expression of relaxin receptors could be controlled by female sex hormones (8), only male rats were chosen in order to minimize the effect of female sex hormone on relaxin of the target tissue. We could not completely control regulating factors associated with relaxin receptor expression, including nutritional conditions, systemic hormonal feedback mechanisms, or the interactions with other hormones such as insulin and glucagon. This study also has some limitations. We found that RXFP1 was distributed only in the transverse carpal ligaments of rats, suggesting that RXFP1 might be a main target for local relaxin administration to treat carpal tunnel syndrome in animal models. We also found that both RXFP1 and RXFP2 were present in the shoulder joint capsules of rats, suggesting that relaxin might be able to loosen fibrotic joint capsules in human patients with adhesive capsulitis. In addition, our immunohistochemical findings failed to reveal whether RXFP1 and RXFP 2 on vascular endothelial cells of inguinal ligament, Achilles tendon, and transverse carpal ligament were involved in regulating the secretion of relaxin. The presence of RXFP1 and RXFP 2 around arterioles suggests that relaxin and the RXFP receptor system might have a role in the regulation of local arterial function as indicated in a previous study (18).
Systemic administration of relaxin has an effect on tissue extracellular matrix components, including collagen and hyaluronan. It can increase the concentration of hyaluronan by upregulating the expression of hyaluronic acid synthetase. Hyaluronan can interfere with the assembly of fibrillar extracellular matrix components, which can results in loosening of the dense collagen fiber network (19). Relaxin has also been shown to have antifibrotic action by downregulating fibroblast activity, increasing collagenase synthesis, and inhibiting transforming-growth-factor–stimulated collagen-1 lattice concentration in rat kidney (20). In addition, relaxin can up-regulate the expression and activities of matrix metalloproteinases (MMPs), a family of extracellular proteases that interfere with matrix remodeling by degrading extracellular matrix components (21). Furthermore, relaxin can downregulate tissue inhibitors of metalloproteinases (TIMPs) (21). Therefore, MMPs can be used to treat fibrotic disease by reducing collagen production and increasing the clearance of TIMPs. Those mechanisms have been explained by systemic administration of relaxin for the treatment of fibrotic diseases. However, local administration of relaxin for musculoskeletal disorders has not been reported yet. Our study is a precedent study on local injection of relaxin for the treatment of stiff ligaments, tendons, and fibrotic joint capsules. Further research is needed to identify the exact mechanisms and interactions between local relaxin hormones and relaxin receptors. Mechanisms of gender-specific responses to relaxin in many target tissues and the value of local administration of relaxin in musculoskeletal tissue remain unclear.
In conclusion, we demonstrated that relaxin receptors RXFP1 and RXFP2 were present in rat musculoskeletal tissues such as ligaments, tendons, and shoulder joint capsules. Studies of relaxin receptors in animal musculoskeletal tissues may provide valuable information for studies on human subjects. In order to understand whether local administration of relaxin is effective and safe treatment for entrapment neuropathies, tendon stiffness, and adhesive capsulitis in humans, animal studies should be preceded. After that, the correct dosage and potential adverse effects of local relaxin injections should be determined before it can be administered to patients in a clinical setting.
 XML Download
XML Download